




הקדמה:
תודתי נתונה לד״ר גדעון רכבי על הערותיו המועילות.

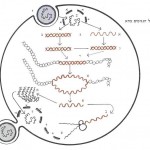
הדנ״א (DNA), כך האמינו הביולוגים מאז פוענח הצופן הגנטי לפני 30 שנה, הוא החומר היציב והשמור ביותר בעולם החי. אפשר להמשיל את גרעין התא לכספת, בתוכה מחזיק האורגניזם את הצופן של כל מגוון החלבונים הדרושים להתפתחותו ולתפקידו. תכניות מוצפנות אלה לעולם אינן יוצאות מתחומי הגרעין. כל פעולות השכפול, התרגום והשיעתוק של החומר הגנטי, מתרחשות בתוככי הגרעין. היחיד שיוצא את תחומי הגרעין אל תוך הציטופלסמה של התא, הוא הרנ״א-שליח (MESSENGER RNA), המהווה מעין תשליל ביו-כימי של הגן הרשום בשפת הדנ״א (תמונה א׳).
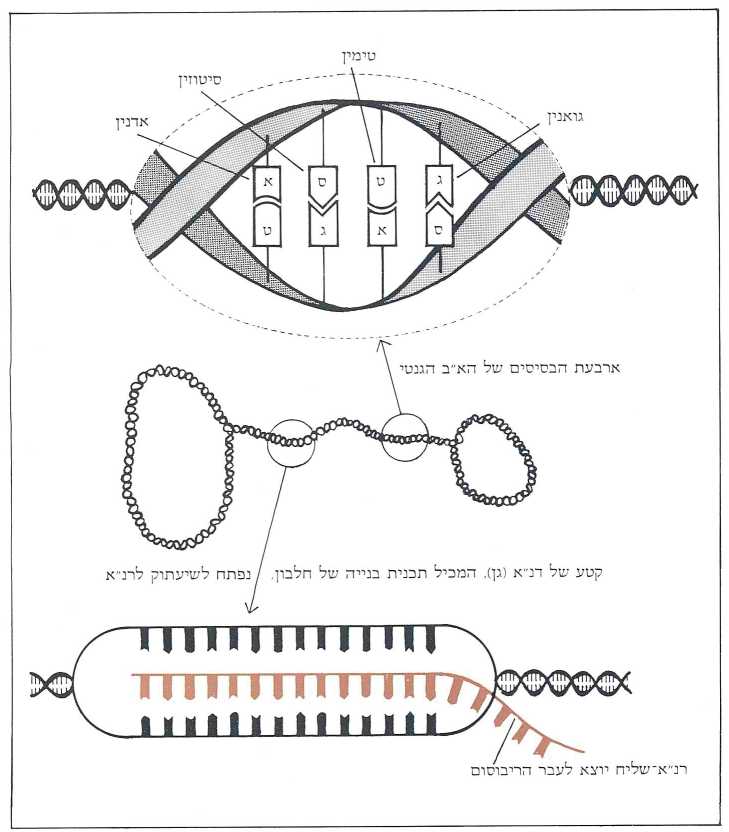
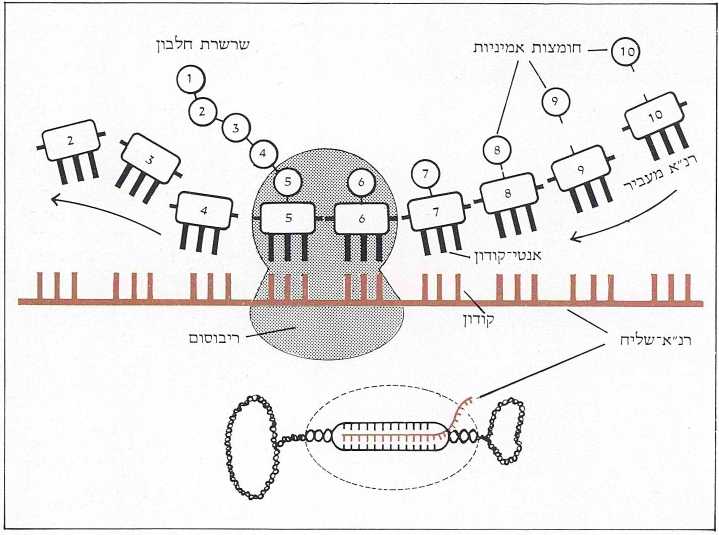
הרנ״א-שליח עושה דרכו לריבוסומים (אתרי ההרכבה של שרשרות החלבון), כשהוא מאורגן בקודונים, דהיינו – במילים בנות שלוש אותיות (בסיסים) מתוך ה-א״ב הגנטי בן ארבע האותיות. בהגיעם לשם הם מתלכדים עם האנטי-קודונים (התואמים אותם) של הרנ״א-מעביר (TRANSFER RNA), שכל אחד מהם צמוד לאחת מתוך עשרים החומצות האמיניות הקיימות. בעזרת רנ״א-ריבוסומי (RIBOSOMAL RNA) נתפרות החומצות האמיניות אחת לרעותה ויוצרות שרשרת חלבונית. רצף זה של חומצות אמיניות תואם את סדר הקודונים של הרנ״א-שליח, ואלה מצדם תואמים בדיוק נחרץ את סדר האותיות הרשומות בדנ״א של הגן. בדרך זאת מכתיב צופן הדנ״א את המבנה של התוצר החלבוני שלו (תמונה ב׳).
אבל, בטבע – כמו בבנקים – אין כספת מושלמת. במהלך מאות מיליוני הפעמים שהדנ״א מתחלק ומשתכפל כדי ליצור תאי-בת, נופלות מעת לעת שגיאות בתהליך ההעתקה. שיבושים אלה עפי״ר מזיקים לאורגניזם ואף עלולים להיות קטלניים; במקרים מסויימים הם נייטרליים, אך במקרים נדירים הם לא רק חיוביים, כי אם יש בהם גם כדי לשאת את בשורת השכלול של10 האבולוציה.

כל כך למה? כי אילו הקפידה האבולוציה לתת לדנ״א הגנה מוחלטת מכל פגיעה אפשרית של קרינות, חומרים כימיים, שיבושי שכפול וכיו״ב, היצורים המפותחים ביותר כיום היו אותם חידקים ואצות קדומים שנוצרו לפני יותר ממיליארד שנה. כדי שייווצרו מנגנונים ביולוגיים יעילים יותר ומינים מפותחים יותר – צריך הדנ״א להשתנות. במילים אחרות, האבולוציה בטבע מתקיימת הודות לשגיאות ולשיבושים המתחוללים בכספת השמורה-היטב של הדנ״א.
תעלומת צבעי התירס
לתוך תמונה נאה וסדורה זאת של האבולוציה, המנווטת ע״י מנגנונים חיצוניים, פרצו בשנים האחרונות ממצאים חדשים שהרעישו את עולם הביולוגיה.
התברר, למשל, שהגנום (המאגר הכולל של הגנים) רחוק מלהדמות לאי רוגע ויציב, המופרע מפעם לפעם ע״י מוטציה. נמצא שגנום זה פרוץ להשפעות ולתמורות וכי הוא מהווה זירה דינמית לתנועה ערה ומתמדת של גנים וקטעי גנים, הנודדים ממקומם הקבוע ומשתכנים באתרים אחרים בכרומוזומים. ושם, במקומם החדש, הם עשויים להעצים או לדכא את פעולתם של הגנים שלצדם התיישבו.
חשד ראשון בדבר קיומם של גנים נודדים התגלה כבר ב-1940 ע״י ברברה מק-קלינטוק, במהלך מחקרה על תורשת הצבעים בגרעיני התירס ואופן התפלגותם בקלחים. מק-קלינטוק ניסתה לעמוד על תופעה מוזרה שלא התיישבה עם כללי התורשה הקלאסיים של הכלאת תכונות. חלק מגרעיני התירס, אף שקיבלו משני ההורים גם יחד גנים לצבע שחור, יצאו אדומים או דו-צבעיים. ברברה מק-קלינטוק הסבירה זאת בנוכחותם של שני גורמים גנטיים (גנים) בעלי תכונות בקרה. האחד שוכן קרוב מאוד לגן של הצבע ואחראי להפעלתו ולשיתוקו לסרוגין, והשני במרחק מה ממנו בכרומוזום – אחראי לוויסות פעולתו של גן הבקרה הראשון. בהעלותה את קיומם של גנים לבקרה, בנוסף לגנים הפונקציונליים (מבניים) שאחראים להפקת חלבונים, הקדימה מק-קלינטוק בעשר שנים את עבודתם של ג’קוב ומונו (FRANCOIS JACOB AND JACQUES MONOD) על פעולתם של גנים המדכאים גנים אחרים.
אבל התגלית שעתידה היתה לזכותה כעבור ארבעים שנה בתהילת עולם, דנה בקיומם ובפעולתם של ״גורמים גנטיים ניידים״ (טרנספוזונים). מק-קלינטוק גרסה כי גן הבקרה יכול להופיע בכרומוזומים של הצאצאים במקום שונה מזה שהיה לו בכרומוזומים של הוריהם ולפיכך גם הגן המבוקר על-ידו הוא אחר. לדעת מק-קלינטוק, נדידת הגנים ממקום למקום אחראית לשוני בביטוי הגנים הצבעוניים בין גרעיני התירס11 בקלח האב לבין גרעיני התירס בקלח הבת (תמונה ג׳).

גילוי זה של גורמים גנטיים ניידים, שהקדים ב-35 שנה את המחקרים אשר הוכיחו את קיומם של ״גנים קופצים״, לא התיישב עם התפישה שרווחה בזמנה בדבר נייחותם וקביעותם של גורמי התורשה. על תגלית זאת זכתה ברברה מק-קלינטוק בפרס וולף (1982) ובפרס נובל לרפואה (1983).
במרוצת השנים התגלו הוכחות לפעולתם של נוודים גנטיים אלה גם בקרב חיידקים ונגיפים. ב-1974 הציעו שני מדענים בריטיים (A. E. JACOB, P. W. HEDGES) הסבר לעמידות המהירה שרוכשים חיידקים מפני תרופות אנטיביוטיות. לדעתם, גנים בעלי עמידות לאנטיביוטיקה מתפשטים במהירות באוכלוסיית החיידקים הודות לכך שהם נישאים ע״י טרנספוזונים.
גילוי הטרנספוזונים פתח צהר חדש להבנת הנתיבים המורכבים של האבולוציה. לדוגמא, קבוצת הגנים לגלובין, הנושאים את תבניות הבנייה של שרשרות החלבון אלפא וביתא של ההמוגלובין (נושא החמצן בכדוריות הדם), ממוקמים באדם בכרומוזומים שונים (11 ו-16). פיצול זה היווה חידה רבתי, עד שהועלתה לאחרונה סברה, לפיה נגרם הפיצול מ״קפיצה״ של גנים. על פי סברה זאת היו הגנים של הגלובין ממוקמים במקורם בכרומוזום אחד. לפני 200 מיליון שנה לערך התחולל רה-ארגון של הכרומוזומים, אשר הביא לפיצולם של הגנים לכרומוזומים נפרדים. הנה כי-כן, בדומה לפיסיקאים, אשר גרעין האטום המוצק והקשיח נתחלף להם בערב-רב של חלקיקים המתרוצצים אלה בתוך אלה, גילו גם הביולוגים כי החומר הגנטי בגרעין התא עשיר יותר במרכיבים ונייד הרבה יותר מכפי שסברו. הגרעין עצמו פרוץ לחידקים ולנגיפים, המעשירים בגנים ובשברי הגנים שלהם את המטען הגנטי של התא. לדעת הביולוג פורד דוליטל, מאוניברסיטת דלהוזי בקנדה, החלה הרבייה המינית בטבע בחילופים כאלה של חומר גנטי בין שני חידקים. כאשר חודר פלסמיד1 של חידק אחד לרעהו, מגיב זה בהוצאת מעין צנור מין, שבאמצעותו הוא מזריק לחידק שכן את הפלסמיד בלוויית חלקים מהדנ״א שלו. הרי לך מהתלה של הטבע: הגרסה הראשונית של הרבייה המינית נושאת אופי מובהק של מחלה מידבקת.
והנה, עוד טרם היה סיפק בידי הביולוגים להסתגל לדיוקנו החדש והדינמי של הגרעין, וכבר הוכו בהלם חדש: הגנום של התא גדוש ומלא אשפה!
אשפת הדנ״א
להפתעתם של הביולוגים התברר, כי תכנית הבנייה של החלבון אינה רשומה בגן באופן רציף אלא במפוצל. האקסונים, קרי: קטעי הדנ״א שמכילים את הקוד של תכניות הבנייה, מפוסקים ע״י רצפי-סרק (אינטרונים), ריקים מקוד ומצופן כלשהו. יתירה מזאת, רצפי האינטרונים ארוכים עד כדי פי-עשר מהאקסונים. וכאילו לא די בכך נמצא גם, כי בין גן אחד למשנהו מצויים רצפים סתמיים של דנ״א, ללא קוד וללא צופן, מעין אשפה חסרת טעם ותכלית. התמונה שנתגלתה לעיני הביולוגים היתה בהחלט מדהימה: שעור ה״אשפה״ בגנום של התא מגיע לכדי 99% מכלל הדנ״א שהוא מכיל. במילים אחרות, אם תכולת המידע של הדנ״א באדם מוערך ב-1700 כרכים בני 1000 עמודים כל אחד, הרי שרק 20 כרכים בהם מודפסים ואילו יתר 1680 הכרכים מכילים עמודים ריקים. והשאלה היא לשם מה ולאיזו מטרה יועדו ספרים ריקים אלה.
שאלה זאת משיאה מייד אצבע מאשימה כלפי הבירור הטבעי: כיצד קרה שהגרזן החד של הבירור הטבעי, אותו גרזן שכורת לאלתר כל שאינו מועיל וכל שאינו יעיל לאורגניזם, לא נחת על אשפה זאת ולא סילק אותה, בעודה באיבה, מעל במת החיים? היו מי שהשיבו, כי עודפי סרק אלה הם דנ״א ״אנוכי״. דהיינו, דנ״א המשתכפל אך ורק למטרה של ריבוי עצמי (על דנ״א אנוכי והתפקיד המיוחס לו באבולוציה של בעלי חיים – ראה ״מחשבות״ 51). ביולוגים אלה, שהבולט בהם הוא פרנסיס קריק (מפצח הקוד הגנטי ושותף לפרס נובל), מניחים, כי במרוצת השכפולים הרבים שהשתכפלו מולקולות הדנ״א במהלך האבולוציה, נוצרו גם רצפים פגומים וסתמיים. על-פי-רוב פגעו שכפולים משובשים אלה בכושר שרידותו של האורגניזם ונכחדו יחד עמו. אבל באותם מקרים שלא היה בהם כדי להזיק ואף לא12 כדי להכביד יתר על המידה, המשיכו להשתכפל ללא מפריע ועברו מדור לדור כנוסעים דוממים ברכב התקין של הדנ״א הפונקציונלי.
הסבר מפתיע ומעניין זה מעלה תמיהה לא קטנה: אפילו אין ברצפים סתמיים אלה כדי להזיק, העובדה שהם מהווים כ-99% מתכולת הגנום, מן ההכרח שיהיה בה הכבדה לא מבוטלת על משאבי האנרגיה של התא, כל אימת שהגנום משתכפל והתא מתחלק. צא וחשוב ש-46 הכרומוזומים השוכנים בגרעין של כל אחד מתאי גופנו (למעט תאי הרבייה) מורכב מ-6 מיליארד זוגות בסיסים (נוקליאוטידים). אם נמתח אותם כחוט ישר יגיע אורכם לשני מטרים. מאחר שגוף האדם מכיל כ-3 ביליון תאים (3×1012) הרי שאורכו הכולל של הדנ״א בגופנו מגיע ל-6 מיליארד ק״מ!! מאחר שכך, נשאלת השאלה מדוע לא פיתח הבירור הטבעי מנגנונים לסילוק פסולת הדנ״א, או לחלופין לא עודד התפתחותם של תאים קלים ויעילים יותר, הנושאים בגרעיניהם רק אותו אחוז בודד של דנ״א פונקציונלי? אבל, אם לא סולק ואם לא נעלם, שמא אין דנ״א סתמי זה אשפה ככלות הכל? שמא שמור לו תפקיד חשוב וחיוני שלפי שעה נעלם מעינינו?
שאלה זאת הביאה את הביולוגים לבחון מקרוב וביתר תשומת לב את ״אשפת הדנ״א״. התברר שאין היא עשויה מקשה אחת וכי מה שנראה ערב רב – יש בו סדר מסויים. נמצא, למשל, כי 5% לערך מעודפי-סרק אלה מורכבים מרצפים קצרים החוזרים על עצמם; 25% מורכבים מרצפים הדירים – אבל ארוכים יותר; 70% הנותרים מורכבים מרצפים חד-פעמיים. התברר עוד שהרצפים הקצרים, בניגוד לבינוניים ולחד-פעמיים, אינם עוברים תרגום לרנ״א, אבל אפשר ששמור להם תפקיד חשוב בוויסות הפעולה של הגנים ואולי גם בשמירת המבנה של הכרומוזומים. ואילו באשר לרצפים האחרים, התברר שמיעוטם מורכב מגנים מדומים (פסאודוגנים), דהיינו גנים שבאחד (או יותר) מבסיסיהם חלה מוטציה ולפיכך נבצר מהם למלא תפקיד פונקציונלי, ובחלקם מורכב מגנים מעובדים בעלי מסלול חיים מוזר ביותר.
המסלול ההפוך של הגן המעובד
מסלול מוזר זה, שנחשף בעקבות התגלית המפתיעה בדבר מבנהו המפוצל של הגן, שינה גם את הידע שהיה לנו על תהליך תרגומו של הדנ״א לרנ״א-שליח. התמונה שנתגלתה הינה ארובה ומורכבת יותר.
כאשר נדרש התא לחלבון מסויים, מועתק הגן (המתאים לו) בשלמותו – על האקסונים והאינטרונים שבו – לרנ״א ראשוני (rna PRECURSOR). בשלב זה נכנסים אנזימים מיוחדים (נוקליאזים וליגזים) לפעולה, החותכים החוצה את האינטרונים ומחברים את קטעי האקסונים זה אחר זה. אנו אומרים חיתוך וחיבור כאילו היה זה דבר של מה בכך, אבל לא הוא. האנזים נדרש לחתוך את רצף הרנ״א בנקודה א׳ ובנקודה ב’ הרחוקה מ-א׳ מרחק של 7,000 בסיסים, ושוב בנקודה ג׳ ו-ד׳ וכן הלאה. במקביל למלאכת הקיטוע נכנס לפעולה האנזים השני, המחבר את קצות האקסונים לפי סדר הופעתם בגן בדייקנות מלאה. ואם נזכור שמרבית הגנים מורכבים מ-20 אקסונים (השיא שייך לגן הקולאגן אלפא המורכב מ-52 קטעים! ), נקבל מושג על דרגת המורכבות של פעולה זאת ועל מידת הדיוק והיעילות הנדרשים לה. מכל מקום, כשמסתיימת פעולת החיתוך והחיבור אנו מקבלים גן מעובד, כעשירית מאורכו של הגן המקורי, המכיל אך ורק את סדר הבסיסים שמייצגים את תכנית הבניה של החלבון הנדרש. גן מעובד זה, או בשמו הרשמי; רנ״א-שליח, יוצא עתה מתחום הגרעין לתוך הציטופלסמה, בדרכו לריבוסומים (תמונה ד׳).
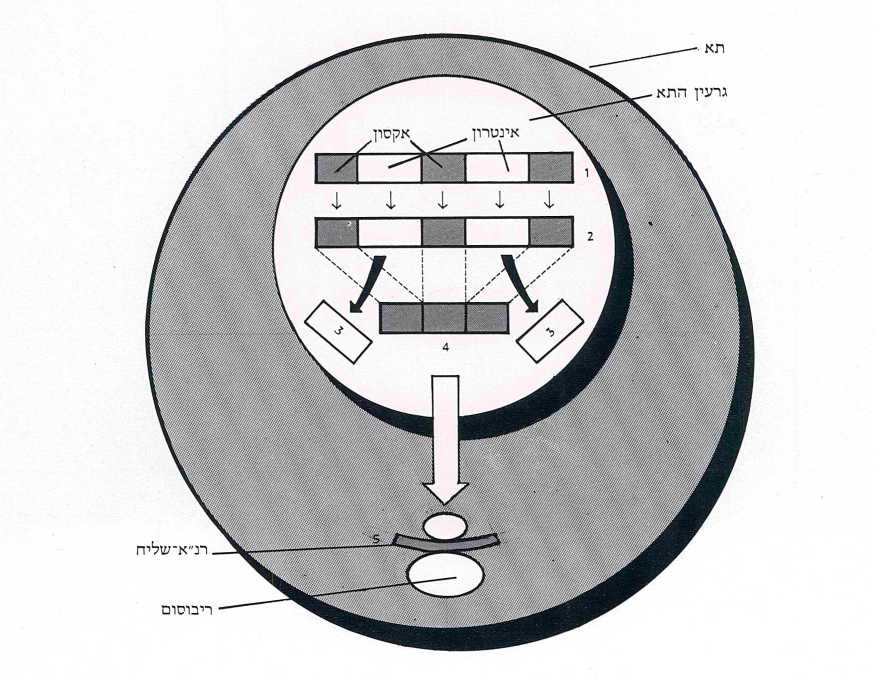
תהליך רב-שלבי ומסורבל זה מעלה ביתר שאת את השאלה בדבר נחיצותם של האינטרונים. שהרי העובדה שנחתכו החוצה ונותרו בתחומי הגרעין, מעידה שאין הם13 נחוצים לעצם התהליך הפיסי של הפקת החלבון בריבוסומים. אם כך, כלום לא היה פשוט ויעיל יותר אילו היה הגן בדנ״א מורכב מלכתחילה רק מאקסונים והיה מיתרגם הישר לרנ״א-שליח, ללא שלב התיווך של הרנ״א-הראשוני? אכן כך, אלא אם-כן הם נחוצים.
הבה נצא איפוא מהנחה שהאינטרונים נחוצים. במקרה כזה מן הדין שהרכב בסיסיהם וסדר הופעתם בכל אינטרון יישמר משכפול לשכפול ומדור לדור, כפי שזה נשמר באקסונים. העקרון ברור: הרכבם וסדר הופעתם של הבסיסים באקסונים נשמר משום שרצף ספציפי זה מייצג במדוייק את תכנית הבנייה של החלבון המסויים, הנדרש שוב ושוב לתא במהלך חייו הוא ובמהלך חיי צאצאיו וצאצאי צאצאיו. אם תחול מוטציה באקסונים או שסדר הופעתם ברצף ישתנה, יחדל התא להפיק חלבון מסויים זה. על-פי אותו הגיון, אם האינטרונים חשובים להפקת החלבון באותה מידה כמו האקסונים, הרי שגם הרכבם צריך להשמר במדוייק. ולהיפך, אם האינטרונים אינם ממלאים תפקיד חיוני, יכולות לחול בהם מוטציות בלי שיעשה נסיון מצד מנגנוני התא לתקנן או לבטלן, שהרי ממילא אין באינטרונים כדי להוסיף או להפחית מחיוניותו של התא. במקרה זה, כל הגנים בקרב בעלי החיים המפיקים אותו חלבון צריכים להיות בעלי אקסונים זהים, אבל יכולים להיות להם אינטרונים בהרכב שונה.
הממצאים בשטח אינם כה חד-משמעיים, כפי שאמנם אפשר להתרשם מבדיקת הגנים לגלובין ולאובאלבומין. האינטרונים מהווים כ-92% מכלל הדנ״א בחמשת הגנים של הגלובין והם שומרים על הרכבם באותה מידה כמו האקסונים (יתרת 8%), הנושאים את תכנית הבניה של הגלובין. ממקרה זה ניתן ללמוד איפוא, שלאינטרונים חשיבות פונקציונלית כמו לאקסונים. לעומת זאת, האינטרונים בגנים של האובאלבומין (חלבון הביצה) נבדלים אלה מאלה אפילו בקרב אוכלוסייה של אפרוחים המורכבת מקרובי משפחה. מסקנה: לאינטרונים אין חשיבות פונקציונלית ביצירת החלבון של האובאלבומין.
ממצאים סותרים אלה הביאו ביולוגים אחדים להציע מוצא של ״פשרה״. עפ״י גרסתם, שינוי באינטרונים בגנים פונקציונליים (גנים מבניים המכילים תכניות בנייה של חלבונים) אין בו כדי לשנות את פעילותם ואת תיפקודם, בעוד ששינוי כזה בגנים של בקרה (הגנים האחראים לוויסות פעולתם של גנים פונקציונליים) אכן משנה את פעילותם.
אם כן, הממצאים אינם מאפשרים להוציא מסקנה חד-משמעית, אך הם מספקים נתון מפתיע ואולי אירוני. מתברר, כי החלקים היחידים באינטרונים, השומרים על הרכב קבוע וזהה בכל הגנים שבקרב כל בעלי החיים, ולו גם ממינים וממשפחות שונות ורחוקות, הם שני רצפים קצרים בשני קצות האינטרונים, המסמנים לאנזימים היכן לקטוע אותם החוצה. במילים אחרות, החלק הקבוע והחשוב באינטרון הוא אותו רצף המביא לסילוקו מתוך הרנ״א-שליח.
שאלת תפקידם של האינטרונים החריפה כאשר גילו בדנ״א של התא גנים רדומים, הזהים בכל לגנים פעילים, בהבדל אחד: הם חסרים אינטרונים. הווי אומר, הם זהים לחלוטין לגנים מעובדים, אותם גנים שעברו תהליך של תרגום לרנ״א-ראשוני ושועתקו לרנ״א-שליח. וכאן נשאלת, כמובן, השאלה, לשם מה נדרש לתא שלב ביניים של העתקת הדנ״א לרנ״א-ראשוני, ואח״כ סילוק האינטרונים מתוכו, כאשר מצויים תחת ידיו גנים ערוכים ומעובדים (ללא אינטרונים), המכילים את כל המידע הדרוש להפקת חלבון ואשר אפשר לתרגמם ישר לרנ״א-שליח.
תשובה בדוקה אין. קיימת רק עובדה מפורשת: גנים מעובדים, בניגוד לגנים רגילים, אינם מפיקים תוצר חלבוני. מעובדה זאת מעריך פרופ׳ דוד גבעול, מהמחלקה לאימונולוגיה כימית במכון ויצמן, שהאינטרונים חיוניים ליציבות הרנ״א במעבר מהגרעין (רנ״א-ראשוני) לציטופלסמה (רנ״א-שליח). אלא שכאן מתעוררת קושיה חדשה: כיצד נוצרו גנים מעובדים בדנ״א? אנו יודעים כיצד מתקבלים גנים מעובדים ברנ״א, בתהליך התרגום של הדנ״א לרנ״א-ראשוני ושיעתוקו של זה לרנ״א-שליח. אנו יודעים גם14 שתהליך זה הוא חד-סטרי, כך קובעת הדוגמה המפורסמת, לפיה זורם המידע מהדנ״א לרנ״א וממנו לחלבונים, אבל לעולם לא בכיוון ההפוך. שאם לא כן, כל שינוי בחלבונים, כגון גידול בשרירים של ספורטאים, היתה עוברת בתורשה לצאצאים. משמע, תורת למארק, אשר גרסה אפשרות של הנחלת תכונות נרכשות של הורים לבנים, היתה נלמדת כיום בבתי-ספר לצד תורתו של דארווין. ואעפי״כ, נוכחות הגנים המעובדים בדנ״א, המהווים העתק מדוייק של הרנ״א-שליח, מעידה לכאורה על קיומה של תנועת מידע הפוכה – לפחות בכל הקשור לחומצות הגרעיניות. דהיינו, מוקדם עדיין להעיר את למארק מקברו, באשר התנועה מהחלבונים לחומצות הגרעין עדיין חסומה, אבל דומה כי תרגום של רנ״א-שליח לדנ״א אכן מתקיים. ואמנם, כאן נכונה לנו הפתעה רבתי: לא זו בלבד שתנועה הפוכה זאת מתקיימת, היא מתבצעת באמצעות נגיפים!
חטיפת הדנ״א
הנגיפים, כידוע, הם טפילים מושלמים. הנגיף מחדיר לתא את החומר הגנטי שלו, משעבד את מערכות הייצור (הריבוסומים) של התא ומאלץ אותו לייצר כפילים בצלמו. התהליך מתנהל בדרך זאת: נגיף אשר החומר הגנטי שלו עשוי מדנ״א, משתלב בגנום של התא ומתחיל להשתכפל כאילו היה דנ״א תאי לכל דבר, בהבדל אחד: קצב שכפולו מהיר הרבה יותר מקצב השכפול של הדנ״א התאי. אותם גנים של הנגיף שנושאים את תכניות הבנייה של מעטפתו, מיתרגמים לרנ״א-שליח ויוצאים מהגרעין לריבוסומים כדי להרכיב שם את חלבוני המעטפת. בתום מלאכת הרכבה זאת מתחברת המעטפת לגנום המשוכפל והשניים יוצאים ארוזים מהתא בנגיף תקין ומושלם – בדמותו ובצלמו של נגיף האב שחדר לתא.
דרכם של הנגיפים שהחומר הגנטי שלהם עשוי מרנ״א (בעל גדיל בודד) – הפוכה וארוכה יותר. הם צריכים תחילה לתרגם את החומר הגנטי שלהם לדנ״א (דו-גדילי) ומכאן שמם: רטרו-נגיפים. התרגום ההפוך, מרנ״א לדנ״א, נעשה בעזרת אנזים מיוחד (רברס-טראנסקריפטאז), שתכניתו רשומה בגנים של הנגיף. בתום מלאכת התרגום של החומר הגנטי של הנגיף, הוא משתלב בגנום של התא ופותח במסלול החיים הרגיל של הדנ״א. דהיינו – משתכפל ועובר תרגום לרנ״א. רצפי הרנ״א שנושאים את תכניות הבנייה של מעטפת הנגיף יוצאים לריבוסומים להרכבת חלבוני המעטפת, שתכליתה לארוז את החומר הגנטי של הנגיף. בכך מגיע תהליך הכפלתו של הרטרו-נגיף לסיומו (תמונה ה׳).
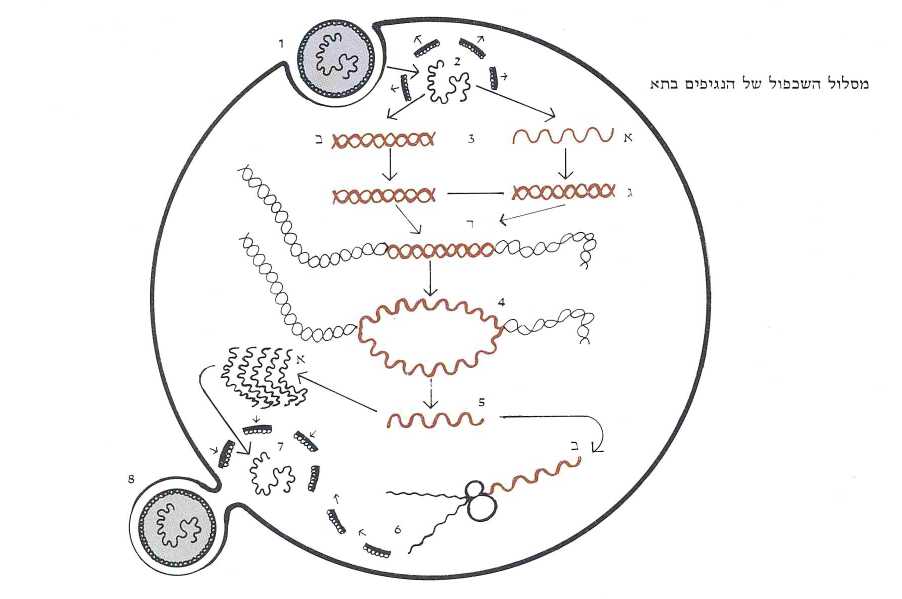
וכאן המקום לשוב לגנים המעובדים שהזכרנו קודם ולשאלה שהצגנו: איך נוצרו הגנים המעובדים וכיצד השתכנו בדנ״א. ובכן, מאחר שהם דומים בכל לגנים התקינים – למעט העובדה שאין להם אינטרונים, ההשערה היא שבמקורם אכן היו גנים נורמליים בעלי אינטרונים. אבל, לאחר שהוצאו האינטרונים מהרנ״א-הראשוני שלהם והם הפכו רנ״א-שליח, נתקלו ברברס-טראנסקריפטאז של העיף, וזה תרגם אותם לדנ״א, כפי שהוא עושה זאת לרנ״א של הרטרו-נגיף. שהרי אין האנזים יכול להבחין בין רנ״א אחד למשנהו, באשר שניהם כתובים באותן אותיות עצמן. וכך, משוכתבים בשפת הדנ״א, שבים גנים נטולי אינטרונים אלה לגנום והופכים להיות גנים מעובדים, או בפי אחרים: רטרו-גנים. ומנין לנו שגנים אלה אכן עברו מסלול רב-תהפוכות זה? בגלל ״טביעת האצבעות״ שהותיר בהם הרנ״א.
מסתבר שבמהלך תרגום הגנים מדנ״א לרנ״א מתווספים בשני קצות המולקולה ראש (CAP) וזנב (poly (A, הבנויים מרצפים של בסיסי גואנין ואדנין בהתאמה2 . והנה נמצא, שהגנים המעובדים בדנ״א מסתיימים בחתימת זנב של (poly (A וברצף בסיסים מוכפל באזור שבו בא אנזים הרברס-טראנסקריפטאז במגע עם הרנ״א. והרי לך טביעת-אצבעות שלא יכול להיות ספק באשר לבעליה.
הנה-כי-כן, גנים אלה, אשר יצאו מהדנ״א עמוסי אקסונים ואינטרונים, שבים עירומים לדנ״א כדי לחיות מרגע זה ואילך חיי נכות, משוללי יכולת לממש את המידע המבני האצור ברצף בסיסיהם.
בכך לא תם עדיין סיפורו של הגן המעובד, ועוד נשוב אליו כדי לגולל את התפקיד הקטלני שממלא אחד מקרוביו. אבל לפני-כן נעשה אתנחתא קלה, כדי לערוך סכום ביניים על האינטרונים ועל אשפת הדנ״א בכלל.
אדם נחות מחידק
אמרנו שמייחסים לאינטרונים תפקיד מרכזי בשמירת מבנה הרנ״א. על כך צריך להוסיף השערה על תפקיד אחר, חשוב מאין כמוהו, שהם ממלאים בקיום האבולוציה ובהאצתה.
משנתגלה שחלוקת הגנים לאקסונים ואינטרונים קיימת רק בבעלי חיים רב-תאיים ולא בחידקים, טבעי היה לראות בגן המפוצל עדות להתפתחות אבולוציונית גבוהה יותר. ככלות הכל, לחידק אין אפילו גרעין, כל הדנ״א שלו מרוכז במעין כרומוזום המרחף בציטופלסמה של התא. לפיכך קיבלו את העדר האינטרונים בגנים של החידק כעדות לנחיתותו האבולוציונית. לא כך חשב פורד דוליטל. הוא הציע להפוך את היוצרות ולראות דווקא בגן המפוצל מצב אבולוציוני ראשוני, פועל יוצא של מנגנון שכפול פרימיטיבי ומרושל שאיפיין את השלבים הראשונים של התפתחות החיים. בעטיו של השכפול המרושל, גורס דוליטל, יצר האורגניזם העתקים רבים של דנ״א, כדי להבטיח שלפחות אחדים בהם יהיו נאמנים ויבטיחו בכך את יציבותו של המידע הגנטי. במרוצת הזמן הפך מנגנון השכפול מדוייק יותר והבירור הטבעי פעל לטובת ההעתקים הנקיים מרצפים סתמיים (קרי: אינטרונים), עד אשר התקבלו גנים רציפים המורכבים רק מדנ״א פונקציונלי (קרי: אקסונים). על-פי תפישה זאת איפוא, הגנים הרציפים של החידקים מייצגים תופעה חדישה ומודרנית יותר מבחינה אבולוציונית מאשר הגנים המפוצלים של האדם. אלא מאי, קיומם של האינטרונים העמיד לרשות הבירור הטבעי מצע רחב ומגוון למשחק המוטציות והצרופים של הבסיסים. מכך יוצא, שדווקא הגנים המפוצלים שימשו מוצא לפיתוח מנגנונים חדשים, ובהמשך גם למינים חדשים.
תאוריה מעניינת זאת אפשר נפגעה מעט לאחרונה, משנמצאו גנים מפוצלים גם בקבוצה מסויימת של חיידקים. אבל, בכל הקשור לפוטנציאל האבולוציוני הטמון באינטרונים, התאוריה בעינה עומדת והיא עולה בקנה אחד עם המודל שהציע ואלטר גילברט מאוניברסיטת הרוורד, בפברואר 1978, בכתב העת NATURE.
גילברט הציע לראות באקסונים ובאינטרונים, במקום בגנים, את שדה הפעולה של האבולוציה. במודל הקלאסי, כאשר מתרחשת מוטציה באחד הבסיסים של הגן, מביא הדבר לשינויה של חומצה אמינית אחת ועקב כך לשינויו של חלבון. מבחינה אבולוציונית זוהי דרך איטית וארוכה ביותר להפקת חלבונים חדשים. ראשית מפני שקצב הצטברות המוטציות האפקטיביות הנו איטי, בערך אחת ל-5 מיליון שנה, שנית – כשמתרחשת כבר מוטציה, הסיכוי שתניב חלבון מועיל לאורגניזם הוא זעיר ביותר. אחת הסיבות היא, שבמודל הקלאסי הגן מהווה יחידה אחת. לפיכך חייב לעבור זמן אבולוציוני רב עד שמתקבל באופן אקראי גן, אשר מלוא הרכב בסיסיו וסדר הופעתם בו יוצרים תכנית מבנה של חלבון מועיל. שכן, אפילו 9,999 מתוך 10,000 בסיסיו רשומים בו בהרכב הנכון ובסדר הנדרש, די שהבסיס האחרון שנותר יהיה שגוי, כדי למנוע מהגן להפיק את החלבון המועיל.
לא כך הדבר במודל של גילברט. מאחר שהגן מורכב מאקסונים, לא צריך להתחיל את כל מלאכת ההרכבה מחדש בגלל מוטציה שגויה אחת. די להחליף את האקסון – שבתוכו מופיעה המוטציה השגויה – באקסון אחר בעל רצף בסיסים תקין. במילים אחרות, מאחר שהמודל של גילברט בנוי על עקרון מודולרי של אקסונים, מתקבלים הגנים המועילים ממשחק צרופים בין האקסונים. יתירה מזאת, כאשר נופלת מוטציה באחד האקסונים, הרי שבהופעתו עם אקסונים אחרים בצרופים שונים, יכול אקסון זה לשמש בסיס ליצירת שורה מגוונת של גנים חדשים. העובדה שהגן מורכב מאקסונים ואינטרונים אף היא חשובה מבחינה אבולוציונית. אם הגן היה רציף ולא מפוצל, אזי אם היינו רוצים לקבל גן חדש מצרוף של אקסונים מגנים שונים, כל אקסון היה צריך להחתך במדוייק במקום הנכון ולהתחבר בדיוק נמרץ בקצה האקסון של הגן השני ואח״כ השלישי וכך הלאה. אולם, כאשר האקסונים מופרדים ע״י אינטרונים, יכול האקסון של גן א׳ להתחבר לאקסון של גן ב׳ בכל מקום שהוא על-פני רצף הבסיסים של האינטרון המחובר לאקסון של גן ב׳.
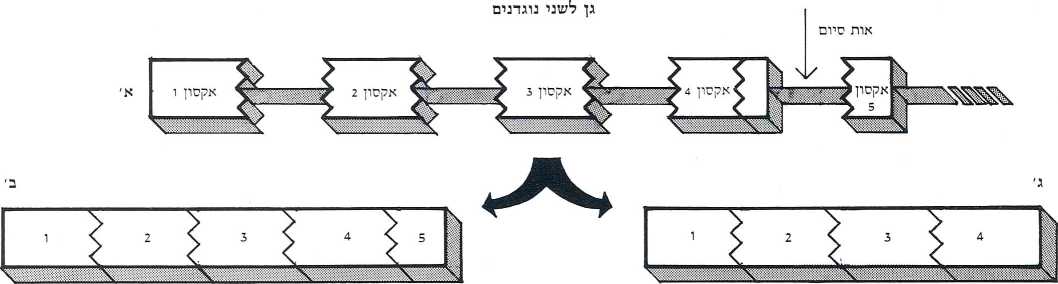
גילברט חישב ומצא שבאמצעות מודל זה אפשר ״להאיץ״ את קצב התפתחותם של חלבונים חדשים בפקטור של 100 מיליון! אפשר איפוא למצוא במודל שלו תשובה אלגנטית לקושי המטריד זה זמן רב את חוקרי האבולוציה והוא, שמנגנון המוטציות הקלאסי יכול אך בדוחק להסביר את 15 הופעתם המהירה של מינים חדשים באבולוציה. זאת ועוד, אפשר להשתמש במודל זה גם כדי להסביר את פעולתם הגמישה והמגוונת של הגנים האחראים להפקת נוגדנים. מספרם של גנים אלה באדם מוערך בכמה מאות. והשאלה היא כיצד הם מצליחים להפיק כמיליון נוגדנים שונים, כנגד מאות אלפי האנטיגנים (גופים זרים) השונים החודרים לגוף, אם אכן גן אחד נושא תכנית של נוגדן אחד? המודלים הקיימים מציעים תשובה ע״י משחק צרופים בין הגנים. המודל של גילברט מציע אף הוא משחק צרופים אך ברמה של אקסונים. המחשה מעניינת לגמישות הטמונה במודל ברמה של אקסונים ניתן לראות בדרך פעולתם של תאי הלימפוציטים מסוג B3 , המייצרים חלבונים של נוגדנים. תאים אלה פועלים בשני שלבים: בשלב ראשון הם מייצרים נוגדנים (מולקולות חלבוניות) בעלי זנב – באמצעותו הם נצמדים לשטח הפנים החיצוני של הלימפוציט, ומעמדה נייחת זאת הם מזהים אנטיגנים. משזיהו נוכחותם של פולשים, מעבירים הנוגדנים הזנבניים מידע עליהם ללימפוציט, והלה מתחיל לייצר כמויות גדולות של נוגדנים, נטולי זנב, המשתחררים לזרם הדם ומשמידים את האנטיגנים.
מכאן, מאחר ששתי הגרסאות של הנוגדן זהות בכל, פרט לזנב, די בגן אחר (בעל חמישה אקסונים) כדי ליצר את שניהם: לצורך הפקת הנוגדנים בעלי הזנב יתבטאו כל חמשת האקסונים ואילו לצורך ייצורם של נטולי הזנב יתבטאו רק ארבעה. האקסון החמישי, המכיל את תכנית הבנייה של חלבון הזנב, מדוכא (תמונה ו’).
גילברט נתן דעתו גם לתפקיד האבולוציוני האפשרי שממלאה אשפת הדנ״א שמחוץ לגנים. אומר פרופ’ גבעול: ״גילברט הציג שאלה מאוד בסיסית לגבי האבולוציה: בפיל יש פי אלף יותר דנ״א מאשר בחידק, איך זה קרה? דרך אחת היא ע״י פעולה מתמדת של צרוף בסיסים. אבל, מדגיש דוד גבעול, ״זוהי דרך טפשית מאוד, מפני שהוספת בסיסים ללא תכנית ובדרך אקראית, היא פסולה לחלוטין. דרך אחרת היא להכפיל כל פעם גנים קיימים. למשל, אם קיים גן שמכתיב חלבון א׳ והוא מוכפל, עומדים בכך לרשות האורגניזם שני גנים זהים שעשויים לשרת שתי מטרות שונות: גן א’ ימשיך למלא את הפונקציה הרגילה שלו, ואילו גן ב׳ – מאחר שאינו דרוש לפעילות השגרתית של האורגניזם – יוכל לעבור מוטציות וליצור חלבון שונה במקצת ממנו. במרוצת הזמן, ככל שגן זה ישתכפל בעשרות ובמאות עותקים, עשוי אחד מהם לקלוט מוטציה וליצור בדרך זאת גן שונה לחלוטין, בעל פונקציה חדשה וחיונית לאורגניזם״.
הנה-כי-כן, אף שחסרות עדיין הוכחות בדוקות וסופיות למודלים אלה, יש בהם כדי להצביע על בך, כי מה שנראה במבט ראשון מיותר וסתמי, מצטייר והולך כנחוץ וחיוני. אמנם, אפשר לשאול מדוע נוירון של יצור פשוט כמו האפליזיה (חלזון ימי), מכיל כמה מאות אלפי פעמים יותר דנ״א מאשר נוירון של יונק, אך מעבר לתמיהות ולשאלות שנותרו עדיין פתוחות, התמונה הכללית מתבהרת והולכת. האינטרונים שוב אינם נתפשים כרצפים בטלים, אלא כמנגנונים בעלי חשיבות אבולוציונית וכמי שמופקדים על שמירת הרנ״א-שליח. הוא הדין ברצפי הדנ״א שמחוץ לגנים. אף שהדיבור על אשפה ועל דנ״א אנוכי מוסיפים להשמע, שוב אין מדובר בגוש אמורפי ונטול סדר. פרט לתפקיד האפשרי שממלאים עודפי דנ״א אלה כבסיס פעולה לבירור הטבעי, חלקם ממלא ככל הנראה תפקיד בוויסות פעולתם של הגנים. אחוז ניכר מ״אשפה״ זאת מורכב מגנים מעובדים, מטרנספוזונים ומפסאודוגנים. פסאודוגן, שמובנו – גן מדומה, הוא גן שחלה בו מוטציה אחת או יותר. הוא מוסיף אמנם לשכון בגנום של התא ולהשתכפל עם הדנ״א הפונקציונלי שבשכנותו, אבל אין הוא מתפקד. קרי: אין הוא מיתרגם לרנ״א ואין הוא מפיק חלבונים. ד״ר גדעון רכבי מבי״ח הדסה ע״ש ״רוקח״ בתל-אביב, מזכיר השערה מעניינת, לפיה הפסאודוגן הוא גן בתהליך מעבר ליצירת גן חדש. מכאן, אפשר שהפסאודוגנים מהווים מאגר בכוח להיווצרות גנים חדשים בתהליך האבולוציה. ואילו הטרנספוזונים, אשר תנועתם בגנום ויכולתם ״לקפוץ״ ממין למין הקנו לחוקי התורשה של מנדל מימד חדש ומרתק, מיקדו לאחרונה את תשומת ליבם של החוקרים, עם שהתברר כי בעוברם ממקום אחד למשנהו בכרומוזום הם עלולים לשמש במקום מושבם החדש כמתגים המחוללים תהליך סרטני בתא.
ומשאמרנו זאת נעבור לחלקה האחרון של רשימה זאת ובה הגילוי המפתיע, כי אנו נושאים בתאי גופנו גנים למחלת הסרטן.
נגיפים מחוללי סרטן
אנו שבים איפוא לרטרו-נגיפים שנזכרו בקשר למעורבותם בהחזרת הגנים המעובדים לגנום. מה שמקנה משנה חשיבות ועניין לחקר הרטרו-נגיפים הוא העובדה שחלק ניכר מהם גורמים להתמרה סרטנית מהירה של התא לתוכו הם חודרים4 .
קיומו של נגיף המחולל סרטן (מסוג סרקומה) בעופות, התגלה כבר ב-1910 ע״י פייטון רוס מהמכון למחקר רפואי ע״ש רוקפלר. אולם, ממצאיו של רוס התקבלו בספקנות רבה והוא נטש את מחקרו. כעבור עשרות שנים הוכח קיומם של נגיפים אלה מעל לכל ספק ורוס זכה בגיל 82 בפרס נובל.16
גילוי נגיפים מחוללי סרטן בחולייתנים הוא ללא ספק ארוע מדעי חשוב, אף שסרטן נגיפי בבני-אדם הוא נדיר מאוד, עד כה נמצא רק רטרו-נגיף אחד החשוד שיש לו קשר משמעותי בהופעת סרטן הלאוקמיה והלימפומה בבני אדם. חשיבותה העיקרית של התגלית, אם-כן, היא בדרך שסללה להבנת מנגנונים מולקולריים תאיים, המעורבים בתופעה הסרטנית.
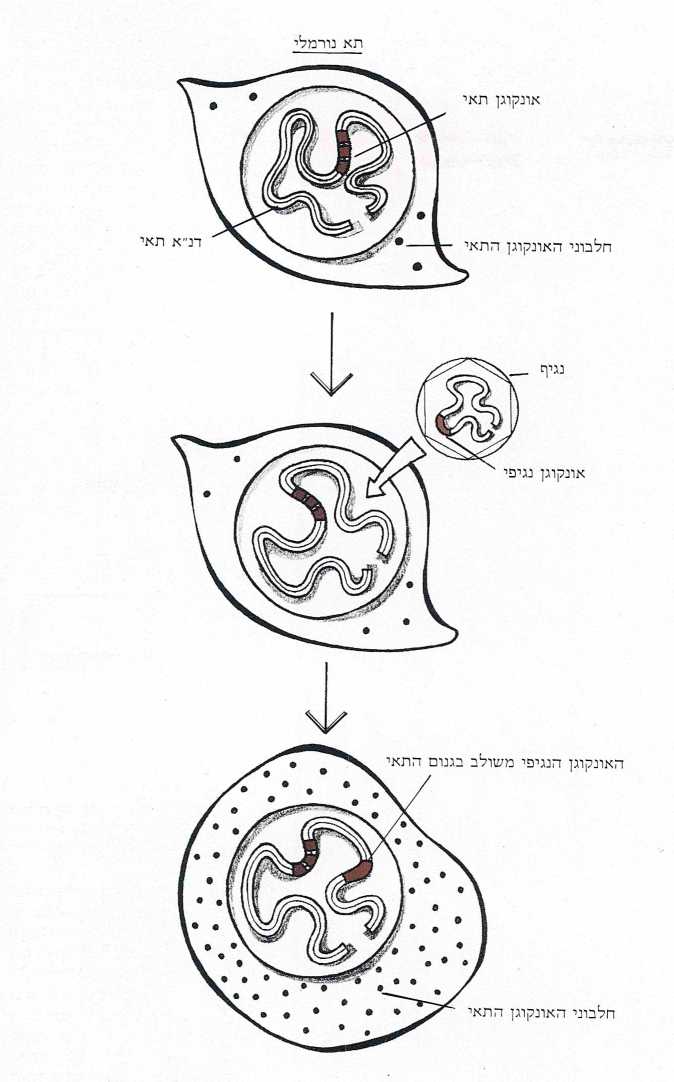
נתחיל בתגלית המפתיעה הראשונה. התברר, שכ-20 מבין הרטרו-נגיפים המוכרים, נושאים בגנום שלהם גן מחולל סרטן (אונקוגן). דהיינו – גן שגורם לתא, לתוכו חדר, להתחלק ללא ריסון.
התגלית השניה, מרעישה אולי אף יותר מקודמתה, הצביעה על כך שמול כל אונקוגן נגיפי מצוי בכל אחד מתאי גופנו אונקוגן תאי, העשוי בדמותו ובצלמו. ההסתברות שדמיון זה הנו מקרי היא קלושה ביותר, לפיכך התעורר חשד, מוזר ככל שיהיה, כי האונקוגנים הנגיפיים אינם אלא העתקיהם של האונקוגנים התאיים. מה שביסס סברה זאת היתה העובדה שלאונקוגנים הנגיפיים אין אינטרונים. מצלצל מוכר? אכן כך. גם לגנים המעובדים השוכנים בגנום של התא אין אינטרונים. ובעצם, למה לא יהיה דמיון? אם נגיף יכול ״לחטוף״ גן שעבר עיבוד לרנ״א-שליח, לתרגמו לדנ״א ולהחזירו לגנום התאי, מדוע לא יוכל ״לצוד״ גן תאי (לאחר שעבר תרגום לרנ״א-שליח) ולשלב אותו בחומר הגנטי שלו, הרשום ממילא בשפת הרנ״א?
ובכן, לא היה מקום לראות ב״ציד״ הגנים יותר מסיפור פיקנטי, אלמלא התוצאות הרות האסון שיש לסיפוח הגן התאי ע״י הנגיף. שכן, בפעם הבאה שיחדור נגיף (מבני קבוצה זאת של 20 רטרו-נגיפים) לתא וישתלב בגנום שלו, יחולל האונקוגן הנגיפי, בעל המוצא התאי, את התהליך הסרטני (תמונה ז’).
עובדה זאת מעלה שאלה מסקרנת: האם השתנה משהו בהרכבו של האונקוגן התאי, במהלך שהותו בגנום הנגיפי, המקנה לו תכונות מסרטנות?
הניסוי של האנאפוזה (HANAFUSA) וצוות עוזריו מאוניברסיטת רוקפלר מזים לכאורה אפשרות זאת. הם הזריקו לעופות רטרו-נגיפים בעלי אונקוגנים פגומים, שהיו חסרים עד שלושה רבעים מכדי תכולתם הגנטית. והנה, תוך זמן קצר השלימו האונקוגנים את החסר להם מתוך החומר הגנטי של התא. בדרך הפכו מבחינת הרכבם לאונקוגנים תאיים לכל דבר, ואעפי״כ שמרו על תכונת הסרטון שלהם. אבל, אם בהרכבם אינם שונים, שמא טמון השוני בצירוף שונה של בסיסים המכתיב לאונקוגן הנגיפי תוצר חלבוני אחר?
ביולי 1983 דווח בעתונות המקצועית על מחקר שערך ד״ר ראסל דוליטל מאוניברסיטת קליפורניה בסן דייגו, באונקוגן (V-SIS) של נגיף ידוע (SSV), הגורם סרטן סרקומה בקופים. המחקר הראה כי מבנה החלבון שמפיק אונקוגן נגיפי זה זהה כמעט לחלוטין לחלבון של גן תאי (PDGF), שתפקידו לגרום להתחלקות מואצת של תאים ברקמות חיבור ושרירים. מסקנה: אם אין הבדל בין תוצריהם החלבוניים של האונקוגן הנגיפי ושל הגן התאי, משמע שהם זהים הן בהרכב בסיסיהם והן בסדר הופעתם במולקולה. ובכל זאת, האונקוגן הנגיפי מחולל סרטן. במה, אם כן, הם שונים?
אפשר שאת התשובה לכך צריך לחפש במקדם (PROMOTER) של האונקוגן. המקדמים הם רצפים קצרים של דנ״א הצמודים לגנים ומשמשים אותם כמתגי הפעלה. מהירות פעולתו של המקדם קובעת את קצב התבטאותו של הגן ולפיכך גם את כמות החלבון שהוא מפיק. בתא נורמלי פועל האונקוגן התאי בהילוך נמוך מאוד, ואילו האונקוגן הנגיפי נמרץ ומהיר לאין שעור יותר. אין זאת שפער המהירות נובע מכך שקיומו של הנגיף תלוי במהירות התרבותו בתא הפונדקאי, ולכן חייבים המקדמים שלו להיות חרוצים ומהירים. ואכן, הניסויים הראו שכאשר מחדירים אונקוגנים תאיים בתרבית תאים – לא קורה דבר. התמרה סרטנית מתחוללת בהם רק כאשר קושרים לאונקוגנים אלה פרומוטרים של נגיף.
אם-כן, עפ״י קו מחשבה זה התופעה הסרטנית נגרמת מהצפת התא בכמות מוגברת של חלבון המופק מהאונקוגן התאי בהשראת המקדם המהיר של האונקוגן הנגיפי.
כך עמדו הדברים עד לפני שנה לערך, כאשר התפרסמו תוצאות מחקר, שיש בהן כדי להעיד כי נוכחותו של האונקוגן הנגיפי איננו תנאי הכרחי להתמרה הסרטנית של התא. החוקרים גילו אונקוגן תאי מסוג מיק-גן (MYC-GENE) בתאי לימפה סרטניים של בני אדם ועכברים, אשר עמיתו הנגיפי לא מעורב בתהליך הסרטני. כיצד, אם כן, הוא מחולל סרטן?
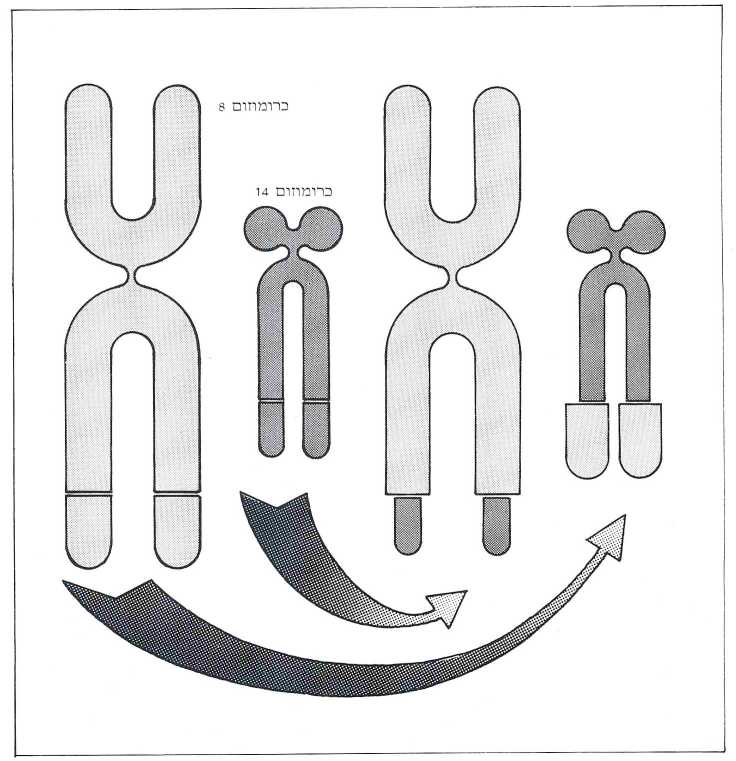
הסברה היא שהמיק-גנים התאיים הם גנים ״קופצים״, המחוללים את התהליך הסרטני בעצם נוכחותם במקום מושבם החדש. מקום מושבו הרגיל של המיק-גן בבני אדם הוא בכרומוזום 8. אבל בקרב 90% מחולי סרטן בלוטות הלימפה מוצאים את המיק-גן בכרומוזום 14 (תמונה ח’). כיצד נעשית קפיצה זאת? נראה כי הדבר מתרחש בעקבות פעולה של שחלוף כרומוזומים בעת התחלקות התא: קטע מכרומוזום 8 מחליף מקום עם קטע מכרומוזום 14. במצב תקין, בשלב ההתמיינות (דיפרנציאציה) של תאי הדם הלבנים, מחליפים מספר כרומוזומים קטעים של דנ״א בתוכם, ״במטרה״ לרכז את הגנים האחראים להפקת נוגדנים, המפוזרים בכרומוזומים שונים, במקום אחד ולגרום להם להפיק את חלבוני הנוגדנים. באדם, למשל, שוכנים גנים אלה בכרומוזומים 2, 14 ו-22. אצל חולי סרטן הלימפה, לעומת זאת, משתבש משהו בתהליך הגורם גם לכרומוזום 8 להשתתף בשיחלוף. מסתבר שהקטע בכרומוזום 8 העובר ל-14 מכיל מיק-גן בדיוק בקו השבר. לדעת פרופ’ דוד גבעול שינוי המקום גורם למיק-גן להתבטאות מואצת ולפיכך להפקת כמויות גדולות של תכנית החלבון שמיק-גן זה נושא בתוכו. מתגי ההפעלה אינם ברורים עדיין. יתכן שהמקדם המהיר של הגנים לנוגדגים מפעיל את המיק-גן שעבר לשכון לידם לפעילות מוגברת, או שהמיק גן משבש את פעולת הוויסות של הגנים לנוגדנים. מחקרים של העת האחרונה שנעשו בעכברים הראו, כי המיק-גן שנודד לכרומוזום 14 מאבד אגב כך את האקסון הראשון שלו. אקסון זה אינו מקדד לחלבון ומכאן מעריכים החוקרים כי תפקידו לבקר את ביטוי המיק-גן, ומשעה שהמיק-גן מתמקם בכרומוזום 14 ללא אקסון הבקרה, הוא מכתיב יצירת חלבון ללא הגבלה. אולם, מה שנכון לגבי עכברים אינו נכון לגבי בני אדם. שכן המיק-גן בתאי הלימפה של האדם עובר בד”כ עם האקסון הראשון – בשלמותו ואעפי״כ מתגלה התופעה הסרטנית.
המסקנה הכללית שמתגבשת היום בקרב החוקרים היא, שפעולת השיחלוף בין הכרומוזומים, המנתקת את האונקוגן מגורמי הבקרה של סביבתו התקינה, גורמת לו לפעול בסביבתו החדשה באורח לא מבוקר, או לחלופין – מנטרלת את מנגנון ההשבתה שלו. כך או כך, דעת הרוב היא שהאונקוגנים מעורבים בכל סוגי הסרטן – בין שהפעלתם נובעת ממוטציה שמתחוללת בהם עקב קרינה או בהשפעת חומר כימי מסרטן או בגין נגיף. זאת ועוד, מתרבים הממצאים המעידים, כי לא די בפעולתו של אונקוגן בודד כדי להפעיל את התהליך הסרטני, אלא בפעולה משולבת של שניים ויותר אונקוגנים. דבר זה יש בו כדי להסביר מדוע לא כל מי שנחשף לגורם מסרטן מפתח סרטן.
מכל מקום, מקרה המיק-גן ודומיו מעלה שאלה מעניינת: האם ההתמרה הסרטנית נובעת מתפוקת-יתר של חלבוני האונקוגנים התאיים, או שמא כל חלבון – אם יציף את התא – עלול לחולל את התהליך הסרטני. במילים אחרות: האם רק האונקוגנים יכולים לחולל סרטן, או כל אחד מעשרות אלפי הגנים הפעילים בגרעין התא מסוגל לכך – אם רק תשוחרר נצרתו בדרך כלשהי?
עפ״י תאוריית ההצפה (OVERDOSE), לא אופיו ולא מהותו של החלבון יוצרים את התהליך הסרטני, אלא כמותו. לדוד גבעול תשובה אחרת. לדעתו, גם גנים אחרים עשויים להיות מופעלים בקצב מוגבר בגין שינויי מקום או שכנות עם גן אחר, אולם יתכן שתוצאותיה של שכנות זאת אינן גלויות לעין. למשל, אם מופעל גן שגורם לבהירות השער, בעל הגן ייעשה בלונדיני יותר. אך מאחר שאין לדבר הזה תוצאות פאתולוגיות כמו למחלת הסרטן, לא נדע שצבע שערו נובע מהפעלת-יתר של גן.
אם נמשיך קו מחשבה זה, אפשר אולי לייחס מחלות שונות לביטוי-יתר של גנים שונים. למשל, מחלת הסכיזופרניה. כבר בסוף שנות ה-60 התברר כי כמות-יתר של דופאמין (אחד החומרים שמשמש להעברת אותות כימיים בין תאי המוח), מעוררת באדם תופעות מובהקות של דמיונות וקולות שווא, האופייניות לסכיזופרניה. העובדה שהמחלה מופיעה בשכיחות גבוהה יותר בקרב בני המשפחה, מחזקת את ההשערה שהיא עוברת ע״י הדבקות נגיפית. אם אכן כך, יתכן שהסכיזופרניה ״איננה״ מחלת נפש, אלא מחלה פיסיולוגית לכל דבר: גן נגיפי מתיישב ליד עמיתו התאי, שאחראי להפקת דופאמין, וגורם לו להגביר את פעולתו.
הנה-כי-כן, אנו ניצבים לפני תמונה חדשה של גנום: פרוץ, נייד ודינמי, אשר במרכזו מתגי-הפעלה גנטיים הקובעים מי, מתי ובאיזה קצב יופעל וידומם כל אחד מבין עשרות אלפי הגנים שבתאינו. מתגים עלומים אלה, הממלאים תפקיד מרכזי בניהול פעילותו התקינה של התא ובשיבושו גם יחד, עשויים להיות מופעלים ע״י גנים של נגיפים שנקלעו באקראי לשכנותם, ע״י גנים שנעתקו ממקומם הקבוע והתיישבו במקומות חדשים,18 
ע”י מוטציות נקודתיות שמתחוללות בגנים, ע״י גנים מעובדים השבים בטעות לגנום, ע״י טרנספוזונים הנעים בחופשיות רבה בגנום ומחוצה לו5 . ומתוך תמונה מורכבת ורבת משתתפים ורבת שמות זאת, מתחיל להתעורר חשד שמא האבחנות שאנו מבחינים בין רצפי סרק שבדנ״א לבין טרנספוזונים6 ולבין הנגיפים, הן מלאכותיות. במילים אחרות: יש יותר מדי מרכיבים שונים בגנום הדומים זה לזה ומתנהגים בצורה דומה מכדי שהדמיון יהיה מקרי. הדנ״א של התא מכיל רצפים פאראזיטיים, נטולי כל תפקיד כשלעצמם, שמשתכפלים עם הגנום של התא. מבחינה זאת הם מתנהגים כנגיפים שהשתלבו בגנום של התא ונחים בתוכו במצב רדום. אבל, אם בעת השכפול ניתק העותק ממקומו ונצמד לאתר אחר בכרומוזום, הרי שהוא נוהג כטרנספוזון לכל דבר או כנגיף לכל דבר. דהיינו, הוא מיתרגם לרנ״א, חוזר ומיתרגם לדנ״א ומשתלב באתר אחר בדנ״א. מאחר שכך, אולי הנגיפים אינם אלא טרנספוזונים, שסיפחו לעצמם קטעי דנ״א ורכשו בכך לעצמם את היכולת לא רק לנוע בתוך הכרומוזום של התא, אלא גם מחוצה לו?
הנה כי כן, כשם שבפיסיקה בוטלה השניות המפורסמת בין גל לחלקיק, אפשר אולי לראות גם את הגנום הביולוגי כאגד של רצפי דנ״א מסוגים שונים, שבתנאים מסויימים הם גנים תקינים ובתנאים שונים ובהרכבים אחרים הם מתנהגים כנגיפים, כטרנספוזונים, כפסאודוגנים וכגנים מעובדים וכיו״ב, ובין מצב למצב הם נחים רדומים כרצפים בטלים וסתמיים של דנ״א.
בדבר אחד שוב אין כמעט ספק. הנגיפים אינם יצורים זרים לגופנו, אלא עצמנו ובשרנו הם, כמו החומר הגנטי העצמי שלנו. הוא הדין בסרטן וברשימה לא קצרה של מחלות לא פחות קטלניות. העיניים לא צריכות להיות מופנות החוצה, לעבר אוייב חיצוני, אלא פנימה, לעבר אותו חומר גנטי רוחש, משתכפל ומונחל מדור לדור – על הנגיפים הרדומים בו ועל גודש הרצפים הסתמיים שבו. ובמשחק הכסאות המוסיקליים הבלתי פוסק שמתנהל בין מיליוני המולקולות הגנטיות הללו, נקבעים חוסנו של האורגניזם וקצב הזדקנותו ונחתך אפילו תאריך היעד של מותו. שכן, כל תא נושא בגרעינו את זרע חורבנו.
ואפשר, כסברתו של ג׳מס גרהם, שהסרטן הוא המחיר שמשלמים בעלי חיים עיליים על טיפוסם במעלות סולם האבולוציה. אם גרהם צודק ונוכחותם של האונקוגנים בתאי הגוף היא מנת חלקם של כל בעלי החיים (למעט יצורים פשוטים כמו הספוגיים), אזי האבולוציה פתחה לפני המינים השונים שתי דרכים להתגונן מפני קרינה אולטרה-סגולית – שהיא מקור סירטון עיקרי: דרכם של חרקים ודומיהם, אשר מימדי גופם הזעיר, קליפתם הכהה ותוחלת חייהם הקצרה מצמצמים הסיכוי להתפתחות התהליך הסרטני, או דרכם של בעלי חיים עיליים. מינים אלה פיתחו מנגנוני שכפול מדוייקים ואנזימים המסוגלים לתקן טעויות שנופלות בעת השכפול. כמו כן הגבילו בהדרגה את יכולת תאיהם להתחדש וויתרו כליל על כושרן של קבוצות תאים מסויימות (תאי עצבים, מוח ושרירים) להתחלק, וזאת כדי למנוע התפשטותן של שגיאות שכפול. מגמה אבולוציונית זאת איפשרה להם לדחות את מועד ההתבגרות ולהאיט את קצב ההזדקנות על-מנת לפתח איברים ומנגנונים מתוחכמים יותר. אבל המחיר, כאמור, היה כבד. הסרטן הוא כרטיס ישיבה במקום טוב באמצע בשורה הראשונה של האבולוציה. האם זהו סוף פסוק? לא. אותה ביולוגיה שמביאה אותנו להבנה קודרת זאת היא גם זאת שנושאת את זרע התקווה. הבנת התהליכים המולקולריים וזיהוי כל האונקוגנים הנעקרים ממקומם בעקבות שחלוף, עשויים לאפשר לנו בעתיד הלא רחוק להתערב במנגנונים המווסתים את ביטוי הגנים, לפתח נוגדנים ודכאנים נגד תאים סרטניים ולעקוף בכך את פעולתו המיכאנית והברוטלית של הבירור הטבעי. בכך לא נעקור את הפוטנציאל הקטלני החבוי בתא כחלק אינטגרלי ממנו, אבל אולי נוכל לעקר את מימושו.
■
צ”י19
טבעת של דנ״א המרחפת ליד החומר הגנטי העיקרי של החידקים ומסוגלת להשתכפל באורח עצמאי. ↩
ההערכה היא כי תפקיד ה״ראש״ לסייע במלאכת התרגום ואילו תפקיד ה״זנב״ להגן על הרנ״א-שליח מפני האנזימים שבציטופלסמה. ↩
מערכת החיסון מכילה גם לימפוציטים מסוג T בעלי תפקיד דומה, אך דרך פעולה שונה משל תאי B ↩
ידועים לפי שעה גם כעשרה נגיפי דנ״א המחוללים סרטן, אבל בתהליך הרבה יותר איטי מאשר נגיפי הרנ״א (רטרו-נגיפים). נגיפי דנ״א אלה משתלבים בגנום של התא ונשארים בו במצב רדום זמן רב יחסית, עד שהם מתעוררים לפעילות סרטנית – אם בהשפעת קרינות וחומרים קרצינוגנים ואם מסיבות אחרות. ↩
קיימת הערכה כי לגן של גלובין פרימיטיבי ולגן של הלגה-מוגלובין (חלבון קטניות שמקבע את החנקן שבאויר), יש מוצא משותף. לפי סברה זאת נויד הגן של הגלובין, במצב מעובד, ע״י נגיף ועבר איתו מעולם החי לעולם הצומח. ↩
לאחרונה נתגלה בצמחים קיומם של נגיפואידים (VIROIDS) זעירים מרנ״א, שגודלם כשליש ממידותיו של הנגיף הקטן ביותר. משערים כי מקורם בטרנספוזונים שאיבדו חלק מתכולתם הגנטית. ↩
