





הקדמה:
מכלל 3.8 מיליארדי השנים שבהן התקיימו חיים על פני כדור הארץ, יותר מ-3 מיליארדים הם היו במצב חד תאי. אף על פי כן, המדע התעלם כמעט כליל מפרק זה באבולוציה, עד שבאה לין מרגוליס. היא הצביעה על כך, שאותו עולם ארכאי התנהל לא על בסיס העקרונות הדארוויניים של תחרות, יריבות ותוקפנות, אלא על יסוד דו קיום, תלות הדדית וסימביוזה. מרגוליס סבורה, שהשערות בחלל הפה והאוזניים, השוטונים של תאי הזרע, הסיבים הקושרים את הכרומסומים בתהליך התחלקותם של התאים, ואולי גם הזנבות הארוכים של תאי העצב במוחנו הם צאצאיהן של בקטריות קדומות שפיתחו דו קיום עם קרבנותיהם או טורפיהם. החיים, אומרת לין מרגוליס, אינם משחק סכום-אפס, שיש בו רק מנצחים ומפסידים.

כשדנים בהופעתן של צורות החיים הראשונות מתייחסים בדרך כלל לאורגניזמים רב תאיים מתקופת הקאמבריון (ר׳ מאמר בגיליון זה על חשיבותה הבלתי נסבלת של המקריות), שהופיעו לפני 570 מיליון שנים ושמהם השתלשלו כל צורות החיים הקיימות איתנו היום. ואמנם, עד שנות ה-50 של המאה הזאת סברו רבים, כי החיים על פני כדור הארץ נוצרו בתקופת הקאמבריון. אבל מאז התגלו מאובנים רבים המעידים על קיומם של אורגניזמים עוד לפני 3.8 מיליארד שנים, ועל כך שהם נותרו בצורתם החד תאית יותר משלושה מיליארד שנים. אם נביא בחשבון שהרב תאים הראשונים הופיעו לפני 700 מיליון שנה, הרי שהחיים החד תאיים תופסים 82% ממשך החיים על פני כדור הארץ. נתון זה צופן בחובו חידה לא קלה: אם למצב הרב תאי יתרונות אנרגטיים מובהקים, תגובה גמישה יותר לתנאי הסביבה ופוטנציאל גדול יותר של גומחות אקולוגיות פנויות, מדוע נדרשו לאבולוציה שלושה מיליארד שנים כדי לפתח את האורגניזמים הרב תאיים הראשונים, בעוד המעבר מיצורים מיקרוסקופיים אלה לדינוזאורים הענקיים ארך רק 500 מיליון שנה?
יתרה מזו, ב-2.4 מיליארד השנים הראשונות, החיים התקיימו כבקטריות ואצות כחולות/ירוקות בעלות מבנה פרוקאריוטי, דהיינו תאים פשוטים ללא גרעין וללא אברונים פנימיים (מיטוכונדריות וכלורופלסטים). ואז, לפני 1.4 מיליארד שנים, הופיעו האיקאריוטים הראשונים, תאים הרבה יותר מורכבים, המאגרים בגרעינם את החומר הגנטי ומכילים אברונים פנימיים.1 אבל למרות מורכבותם נדרשו 700 מיליון שנה ויותר עד שהתפתחו מהם היצורים הרב תאיים הראשונים. שאלות פתוחות אלו חשובות להבנת האבולוציה, אך ספק אם הן היו מתעוררות כלל אלמלא לין מרגוליס, פרופסור למיקרוביולוגיה באוניברסיטת מסצ׳וסטס, אשר רעיונותיה הנועזים ומאמציה הבלתי נלאים לחשוף את עולמם המרתק של המיקרואורגניזמים העמידו את החד תאים במוקד העניין המדעי. יתרה מזו, עבודתה של מרגוליס מעלה קושיה יסודית לגבי המכניזם של האבולוציה: האם הוא מודרך על ידי אגואיזם צרוף ומאבק אלים – כגירסת הניאו דארוויניסטים – או על ידי עיקרון של תמורה הדדית ושיתוף פעולה?
מאז טבע הפילוסוף הבריטי הרברט ספנסר (1903-1820) את הביטוי המפורסם (שבטעות מיוחס לדארווין) שרידות הכשירים (Survival of the Fittest), נוהגים לייחס לאבולוציה הדארווינית את העיקרון של מלחמת הכל בכל. על פי עיקרון מיליטנטי זה, הברירה הטבעית מעודדת אלימות, כוח, תוקפנות ואגואיזם, כיוון שתכונות אלו מאפשרות לפרט להשיג מזון, להגן על נחלתו, להפרות יותר נקבות ולהבטיח בכך תפוצה רבה יותר לגנים שלו. תפיסה זו, שהתוקפנות והאלימות טבועות בגנים שלנו, הניבה שפע מסקנות פילוסופיות ומשנות חברתיות, החל בפרי הבאושים של התורה הנאצית, עבור בתזות שנויות במחלוקת נוסח התוקפנות המובנית של קונרד לורנץ ומורשת הקוף הרצחני של ריימונד דארט, וכלה בברירת קרובים של הולדיין והמילטון, בסוציוביולוגיה של וילסון ובגן האנוכי של דאוקין. והנה באה מרגוליס ומצביעה על הפכיהן של תכונות אלו – שתפנות, תלות הדדית וסימביוזה – כעל המנגנונים המרכזיים בשלושת מיליארד השנים הראשונות לקיום החיים, שמשקלם האבולוציוני רב גם היום.
העצמי והלא עצמי
שורשי התפיסה הקואופרטיבית של מרגוליס נעוצים בעבודת הדוקטור שלה משנת 1965, לפיה אברונים מרכזיים בתאי החי והצומח, כמו המיטוכונדריות המספקות אנרגיה לגוף והכלורופלסטים המעורבים בתהליך הפוטוסינתזה, הן בקטריות קדומות שנבלעו על ידי תאים איקאריוטים ונטמעו בתוכם. התזה הפרובוקטיבית של הסטודנטית הצעירה עוררה לעג בקהילה המדעית, אך כיום איש לא חולק עליה. בהמשך יצאה מרגוליס במסע יחיד למיקרוסמוס, כדי לעמוד על חשיבותם של היצורים החד תאיים לאבולוציה בכלל ולקיום החיים המאקרוסקופיים בפרט.
מסתבר כי במהלך שני מיליארד השנים הראשונות לקיום החיים על כדור הארץ פיתחו הפרוקאריוטים מנגנונים כימיים מתוחכמים ליצירת תסיסה, פוטוסינתזה ונשימת חמצן. מנגנונים אלה ואחרים איפשרו לבקטריות לקשור ישירות חנקן מהאוויר, לשנות את הרכב האטמוספירה ולגרום לתמורה יסודית בקרום החיצוני של קליפת כדור הארץ. חשיבותם של החד תאים אינה רק היסטורית. בלעדיהם אין להבין את תיפקוד המערכות האורגניות המודרניות של יצורים מאקרוסקופיים, ובמידה רבה גם את הגיאוכימיה של כדור הארץ. לדוגמה, הבקטריות ממלאות תפקיד מרכזי בטיהור מאגרים של מי שתייה ובחידוש פוריות האדמה; הן מייצרות מזון ואנרגיה מפגרים וצמחים רקובים ומעכלות את התאית הקשה בבטנם של טרמיטים ואוכלי עשב; צמחים ממשפחת הקטניות לא היו יכולים לקשור את החנקן במישרין מהאוויר אלמלא שיתוף הפעולה שלהם עם בקטריות החיות בשורשיהם; העובר האנושי מתפתח בסביבה נוזלית שמליחותה דומה לזו של האוקיינוסים הקדומים, וגם ריכוזי האשלגן, הנתרן והכלור ברקמות שלנו כמעט זהים לאלה של הסביבה הקדומה. ולבסוף, גוף האדם 20מכיל מאה טריליון (1017) תאים שהתפתחו מבקטריות, המהווים 10% ממשקלו היבש.
האם האבולוציה מודרכת על ידי אגואיזם צרוף ומאבק אלים או על ידי תמורה הדדית ושיתוף פעולה?
ובכן, כיצד נוצר שיתוף פעולה זה וכמה זמן היה דרוש לו כדי להתפתח? לפני 5 מיליארד שנים לערך החל ענן אדיר של גז ואבק, שנפלט לחלל מהתפוצצות של כוכב ענק (סופרנובה) באזורנו, לעבור דחיסה גרביטציונית, אשר הציתה בו בערה גרעינית. כך נולדה ככל הנראה השמש שלנו. לא עבר זמן רב, ושטף הקרינה שהתפשט ממנה עורר לחיים גם את האטמוספירה הדלה שהקיפה את גוש החומר ממנו התפתח כוכב הלכת שלנו. גיל כדור הארץ הוא קרוב לוודאי 4,600 מיליון שנה, אך ב־700 מיליון השנים הראשונות לקיומו הוא לא גילה מידה של הכנסת אורחים. התפרקויות רדיואקטיביות אינטנסיביות של אורניום, תוריום ואשלגן בגרעינו עשוהו כדור לבה לוהט ומנעו היווצרותה של קליפת חומר קשה. מים שנוצרו מהתרכבות של חמצן עם מימן התפרצו מפנים כדור הארץ בזרמי קיטור אדירים והתערבבו באטמוספירה עם דו תחמוצת הפחמן ועם גזים רעילים של ציאניד ופורמלדהיד. אבל לפני 3.9 מיליארד שנה התקררו פני האדמה במידה מספקת כדי ליצור קרום יבשתי דק ולאפשר לאדי הקיטור להתעבות לטיפות מים. גשם זלעפות, שנמשך ללא הפוגה כמה אלפי שנים, שטף את הסלעים ממלחיהם וסחף אותם לימות ולאוקיינוסים. במקביל התקיימה פעילות וולקנית עזה, שפלטה לאוויר גזים מבטן כדור הארץ, ואלה העשירו את האטמוספירה ואת הימות בתרכובות כימיות חדשות. התנאים הכימיים והפיסיקליים הבשילו עד מהרה להופעת החיים, ודי היה בניצוץ ברק, בהדף רעם, בקרינה אולטרה סגולה או אפילו בחום בלבד כדי ליצור את אבני הבניין של החיים: חומצות אמיניות וחומצות גרעיניות.

על הופעתם הספונטנית של החיים, הכרוכה ביצירת אנזימים ומולקולות רנ״א בעלות כושר שיכפול עצמי, עדיין לא נאמרה המלה האחרונה ואנו עוד רחוקים מהבנתה. אלא שלא בסוגיה זו עוסקת רשימה זו (על היווצרות החיים, ר׳ מאמריהם של מישל רבל, ישעיהו ליבוביץ ושניאור ליפסון, מחשבות 34-33, 35 ו-57), כי אם באירועים העלומים שהתרחשו 100 מיליון שנה אחרי הופעת החיים. אז התחילו מולקולות של חומצות גרעיניות להתעטף בקרומיות שומניות, ובכך יצרו לראשונה את האבחנה החשובה מאין כמוה בין העולם החיצוני לעולם הפנימי, בין העצמי ללא עצמי. שלב זה מבשר את הופעת החד תאים, להם מקדישה לין מרגוליס את ספרה Microcosmos2 .
בשורת האור
המעטפת הקרומית היתה קריטית לקיומן ולתיפקודן של המולקולות האורגניות הקדומות. בחסותה יכלו חומצות הגרעין (הדנ״א והרנ״א) לשכפל את עצמן בביטחון יחסי, ולהעביר לתאי הבת תכונות מועילות שרכשו באקראי; הקרומית חסמה כניסתם של חומרים עוינים לתא, ומצד שני איפשרה לחומרים חיוניים לחדור לתוכו. סלקטיביות זו היתה חיונית ביותר לבקטריות הראשונות, כי לא היה להן כנראה די חומר גנטי לקידוד כל החלבונים הדרושים, ועל כן נאלצו לקלוט אותם במישרין מסביבתן העשירה בתרכובות ביוכימיות. אלא שארוחות חינם אלו הלכו ואזלו ככל שאוכלוסיית הבקטריות גדלה, וזו אכן התרבתה בקצב מדהים. בקטריה זריזה מסוגלת להתחלק כל עשרים דקות; בשתי יממות יכולים להיווצר אפוא 2144 פרטים, יותר מכל בני האדם שחיו אי פעם; בארבעה ימים עשויה אוכלוסיית הבקטריות למנות 2288 פרטים, יותר ממספר הפרוטונים (2266) ביקום.
כמובן, רק חלק זעיר ממספר אסטרונומי זה מתממש. רובו המכריע נכחד בידי הב21רירה הטבעית, המעמידה במבחן מעשי את המוטציות האקראיות, פורסת חסותה על היעילות והחיוביות יותר, ובכך מעניקה כיוון לאבולוציה. קצב המוטציות בחומר הגנטי של הבקטריות אינו עולה על מוטציה אחת למיליון חלוקות, אבל סקאלת הזמן של האבולוציה היא גדולה דיה כדי לאפשר הופעתם של חידושים חיוביים. אחד החידושים הללו התבטא בהתפתחותו של מנגנון תסיסה, המפיק מולקולות עתירות אנרגיה מסוג ATP. מנגנון התסיסה איפשר לבקטריות לסנתז תרכובות מזון בעודן חבויות באדמת הביצות, מוגנות מפני הקרינה האולטרה סגולה הקטלנית של השמש. פתרון זה היה כה מוצלח, עד כי מנגנוני תסיסה דומים מוסיפים לשמש גם בימינו אורגניזמים שונים, לרבות אותנו. למשל, במצבים של מאמץ גופני מתמשך, כאשר אספקת החמצן אינה עומדת בדרישה הגוברת לאנרגיה, מפעילים תאי השריר מטבוליזם רזרבי של תסיסה.
עם זאת, תהליך התסיסה אינו מצטיין ביעילות אנרגטית. על כן, הופעת מנגנון הפוטוסינתזה היתה אירוע בעל חשיבות עליונה בתולדות החיים על פני כדור הארץ. יש להניח, כי בשלב ראשון נטלו הבקטריות את המימן ואת דו תחמוצת הפחמן הישר מהאוויר, כדי ליצור באמצעותם את התרכובות הפחמניות הנחוצות למזון ולבניית חומרי התא. אולם רזרבות המימן החופשי אזלו במהירות מהאטמוספירה, כיוון שכוח הכבידה של כדור הארץ לא היה חזק דיו לעצור בעד המימן הקל מלברוח לחלל החיצון. לחץ הברירה הטבעית התחיל לפעול אפוא לטובת בקטריות שניצלו את אנרגיית האור ליצירת מולקולות ATP, אשר סיפקו להן את האנרגיה הדרושה לחילוץ המימן מהסולפידים שפלטו הרי הגעש לאוויר. אבל גם הפעילות הוולקנית האינטנסיבית לא יכלה לעמוד לאורך זמן בביקוש הרב למימן. את הפתרון לבעיה זו סיפקה בקטריה קטנה (סיאנובקטריה), אשר מוטציה מוצלחת שחלה בה הוסיפה למנגנון הפוטוסינתזה הסולפידי מוקד פוטוסינתטי שני. מוקד זה העצים את האנרגיה של המנגנון הראשון ואיפשר למערכת הפוטוסינתטית לפרק את הקשר הכימי החזק בין המימן לחמצן במולקולת המים. בכך לא רק נפתח לפני הסיאנובקטריה מקור אינסופי של אנרגיה, כי אם גם ניתן בידה מנגנון אנרגטי יעיל פי 18 מתהליך התסיסה.
הסיאנובקטריות הוכיחו עצמן כלהיט אבולוציוני מרגע הופעתן. מצוידות בשני המוקדים הפוטוסינתטיים, הן התפשטו ללא הפרעה וכיסו במעטה דק וירוק כל אזור רטוב ומואר. הצלחתן רשומה עד היום בסטרומטוליטים, אותם סלעים עגולים המעטרים את החופים של איי בהמה ומערב אוסטרליה, המורכבים משכבות של סיאנובקטריות מתות – זו גל גבי זו – בדומה לשוניות של אלמוגים. ניתן לראות גם היום את המעטה הירקרק-חום שלהן על סלעים רטובים, בבריכות שחייה ועל וילונות האמבטיה. אולם פעילותן המוצלחת היא רק חלק אחד בסיפור, שכן על כל אטום מימן שהן הפיקו ממולקולות המים, השתחררו לאוויר שני אטומים של חמצן. אמנם בימינו החמצן הוא סם חיים, אבל עבור העולם החד תאי הקדום הוא היה גז רעיל מאין כמוהו, משום נטייתו לחמצן שומנים, חלבונים, סוכרים וחומצות גרעיניות, ולגרום בכך לפירוקם. אגב, גם תאי גופנו מצוידים במנגנונים מיוחדים שנועדו להגן עליהם מפני חימצון יתר. יתר על כן, למרות שני מיליארד שנות חיים עם חמצן יחסנו אליו עדיין אמביוולנטי, ולא בכדי. מחקר חדש יחסית מראה שאורך חייהם של זבובים קצוצי כנפיים גדול פי 2.6 מזבובים מכונפים. הסיבה המשוערת היא, שתוספת החמצן שצורכים בעלי הכנפיים בעת מעופם הופך להידרוקסיל (OH), המחמצן את חומרי התא ומאיץ את תהליכי ההזדקנות.
מעוינות לשותפות
ובכן, ככל שהצלחת הסיאנובקטריות גברה, כך הלך והזדהם האוויר בחמצן, עד שלבסוף סיכן את החיים עלי אדמות. השואה הבלתי נמנעת היתה כתובה על הקיר באותיות מחומצנות. ואמנם, אוכלוסיות שלמות נשמדו, למעט כמה בני מזל שהצליחו להתגונן מהחמצן באמצעות ויטמין E, וכמובן אותן בקטריות שהקדימו למצוא מסתור מהקרינה האולטרה סגולה בביצות האנאירוביות ובמעיינות חמים של מימן גופריתי. ייתכן שהחיים היו נעצרים שם, אלמלא התמזל מזלן של כמה סיאנובקטריות לעבור סדרה מוצלחת של מוטציות, שבסופה נוצר מנגנון חדש (מיטוכונדריה), המסוגל להפיק אנרגיה בתהליך של נשימת חמצן. זן חדש זה של סיאנובקטריות היה מצויד אפוא בשני מנגנונים משלימים: מנגנון פוטוסינתטי המספק מימן לבניית פחמימות, ומנגנון מיטוכונדרי, המשתמש בחמצן הרעיל שפולט המנגנון הראשון כדי להפיק דו תחמוצת הפחמן, מים ושפע אנרגיה. למעשה, המיטוכונדריה היא תחנת אנרגיה משוכללת, המסוגלת להפיק 36 מולקולות ATP על כל מולקולת סוכר שהיא מפרקת, כפול ממנת האנרגיה שמפיק תהליך הפוטוסינתזה. לסיאנובקטריה החדשה היו אם כן כל הכלים הדרושים לפיתוח מסלול אבולוציוני ארוך ומשגשג. היא היתה מסוגלת לספק את צרכיה בתנאי אור וחושך גם יחד, בלי שתיאלץ להתחבא בבוץ מפני הזיהום החמצני. על הצלחתו של מסלול זה אמרה ההיסטוריה את דברה: נושאי המיטוכונדריות בימינו הם אריות, פילים ובני אדם, ואילו צאצאי הבקטריות 22שמצאו בשעתו מסתור בביצות חיים היום במעיהם.
הבקטריות שהיו לפנים אויבן הקטלני של האמבות התמזגו עימן.
תפוצתם המהירה של הסיאנובקטריות ושל מיקרואורגניזמים אחרים שהלכו בעקבותיהן, חוללה שינויים עמוקים באטמוספירה. מהרכב מקורב של 98% דו תחמוצת הפחמן, אפס חמצן ו-2% חנקן לפני הופעת החיים, התייצבה האטמוספירה על 0.03% דו תחמוצת הפחמן, 21% חמצן ו-78% חנקן (וכן שאריות גזים אחרים). מעבר לכך, תרומתה המשמעותית ביותר של הסיאנובקטריה להתפתחות החיים על פני כדור הארץ באה לידי ביטוי בסלילת הדרך להופעת הפרוטיסטים, קרי תאים איקאריוטיים בעלי אברונים פנימיים וגרעין מרכזי עתיר דנ״א, שבלעדיהם לא היה נוצר עולם החי והצומח המוכר לנו היום.
העובדה שהכלורופלסטים והמיטוכונדריות מכילים דנ״א ורנ״א משל עצמם, מאפשרת להם לייצר את החלבונים הדרושים להם, ללא תלות כמעט בדנ״א המרכזי של התא. עצמאותם שיכנעה את מרגוליס שהכלורופלסטים והמיטוכונדריות לא התפתחו בסיאנובקטריה בתהליך הדארוויני הרגיל של מוטציות מקריות, כמתואר לעיל. לדעתה, אברונים אלה היו בשעתם בקטריות עצמאיות, שנבלעו על יד הסיאנובקטריה ופיתחו במרוצת הזמן סימביוזה מלאה איתה. על פי תסריט אחר, אבי המיטוכונדריות היה טורף מיקרוסקופי, שנהג לפלוש לגופן של סיאנובקטריות ולעכל אותן מבפנים. סביר להניח, שרוב הסיאנובקטריות נעכלו על ידי המיטוכונדריות שפלשו אליהן, אבל כפי שקורה לא אחת לחיידקים ונגיפים אלימים (ר׳ סיפורו של קוונג להלן), תוקפנות יתר לא משתלמת, משום שחיסול הפונדקאי מביא גם למות הפולש. בלחץ הברירה הטבעית התפתח אפוא זן מתון יותר של מיטוכונדריות, שלא עיכל את הסיאנובקטריה, אלא השתכן בתוכה, ניזון מחומריה ומאשפתה, ובתמורה סיפק לה אנרגיה. הברירה הטבעית לא איחרה לגלות את היתרון שבשיתוף פעולה זה, והיא עודדה את המשך הניסוי, עד שהתגבשו יחסים סימביוטיים מלאים בין שני האורגניזמים. לימים נבלע יצור סימביוטי זה על ידי תא איקאריוטי, אשר הותיר את 23המיטוכונדריה בשלמותה ויצר עם מי שהיתה לפנים טורף, ועתה הפכה טרף, מערכת סימביוטית דומה. תסריט זה של דו קיום רב שלבי משמש לדעת מרגוליס עדות ליתרון האסטרטגיה של שיתוף פעולה על פני תחרות ואלימות.
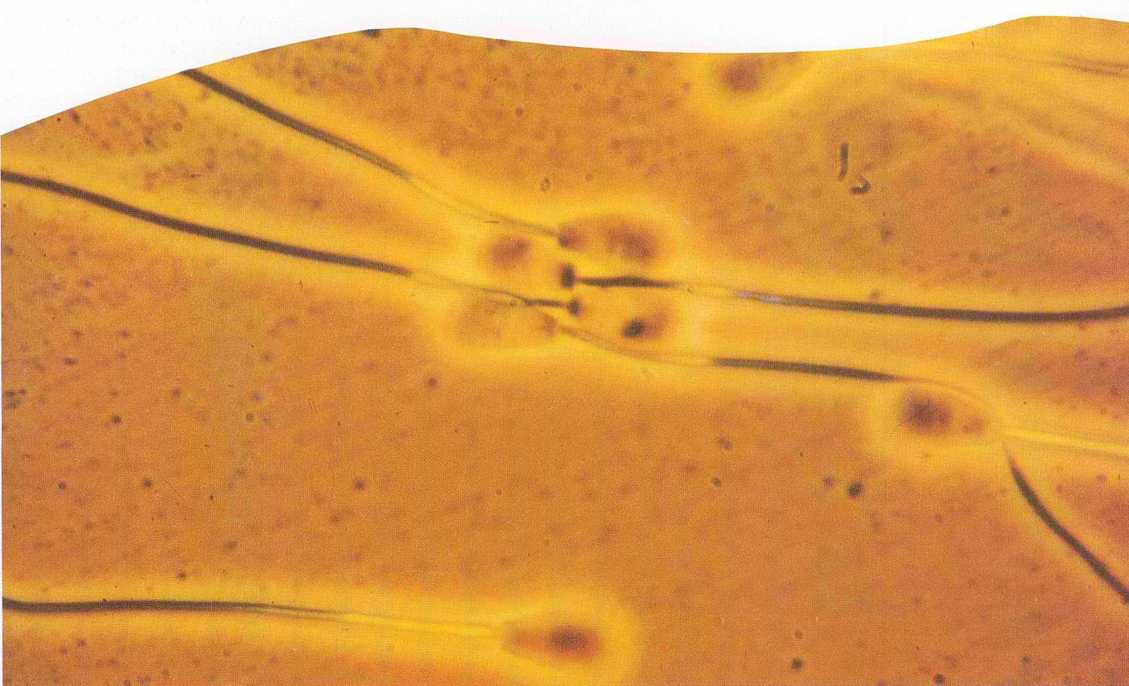
תודעה בקטריאלית
לא ארכו הימים ומרגוליס הצביעה גם על השוטונים והשערות של איקאריוטים שונים, המשמשים אותם לתנועה, כעל מי שהיו לפנים בקטריות עצמאיות, מסוג ספירוכטים דמויי שוטון, שוויתרו על עצמאותם ועל ניידותם תמורת יחסים סימביוטיים עם פונדקאיהם. בשלב כלשהו עלה בדעתה, שגם השערות בחלל האף והאוזניים שלנו, וכך גם השוטון של תאי הזרע, הם אולי צאצאי אותם ספירוכטים ניידים. הערכה זו התחזקה נוכח העובדה, שלשערות האוזן והאף, לשוטוני הזרע, לסיבי הכישור בתהליך התחלקות התא ולספירוכטים כמה תכונות משותפות: מבנה צינורי בסיסי, הרכב החלבונים, העדר כושר התחלקות ומערך חיסוני דומה. כמובן, אין די בדמיון זה משום אישור לתזה של מרגוליס. אף על פי כן, יש משהו מדהים בעובדה שאורך הדנ״א בכל תאי גופנו, הגדול פי מאה מיליון מהמרחק בינינו לירח, מתפקד בתיאום מושלם כל כך – למרות היותו תאגיד של אורגניזמים.
אך דומה כי האפשרות שאורגניזמים בקטריאליים אלה מצויים גם במוחנו מרעישה עוד יותר. הייתכן שהדנדריטים הדקים והאקסונים הארוכים של תאי המוח היו אף הם אי פעם ספירוכטים? גם אם השערה זו לא ניתנת לאישור, יש בה כדי להלהיב את הדמיון. צא וחשוב, גופנו בנוי מתאגיד ענק של איקאריוטים ובקטריות, המקיימים אורח חיים שיתופי כמו בקומונה. גם תאי מוחנו, שבאמצעותם אנו מנסים לעכל רעיון פנטסטי זה, הם כימרות (מפלצות מיתולוגיות) שנוצרו ממיזוג קדום בין תאים פרוקאריוטים ואיקאריוטים. יוצא אפוא, שהתודעה היא פועל יוצא של עיקרון סימביוטי.
רעב שהוליד את המין
העיקרון הסימביוטי חל גם על מערכת הרבייה בטבע. במרוצת 50 השנים האחרונות גילו מדענים, שהבקטריות מחליפות ביניהן קטעים של חומר גנטי. פיסות אלו של דנ״א, המכונות רפליקונים, מעבירות מידע בין הבקטריות ומשמשות לתיקון קרעים בדנ״א. בעצם, אם נגדיר את הפעילות המינית כהחלפת חומר גנטי בין שני פרטים, נוכל לראות בחילופי הרפ-ליקונים מגעים מיניים קדומים, המתנהלים באינטנסיביות רבה ובמתירנות מלאה. אמנם הרבייה המינית התפתחה לפני שני מיליארד שנה לערך, בעוד הבקטריות מתרבות זה שלושה מיליארד שנה ברבייה לא מינית, על ידי חלוקת התא לשני 24תאי בת (שכל אחד מהם זהה לתא האם), אבל מכניזם זה של חילופי רפליקונים הביא את מרגוליס להרהר מחדש במוצא הרבייה המינית ובסיבות האבולוציוניות להתפתחותה.
הרבייה המינית כרוכה בתהליך מייגע ומסובך של מציאת בת/בן זוג, ביזבוז אנרגיה על מאבק במתחרים ועל טקסי חיזור, הסתכנות במגע גופני עם פרט זר, והרפיית הכוננות מפני טורפים מזדמנים בעת האקט המיני. יתרה מזו, כדי להביא תהליך זה לסיומו המוצלח נדרשה הברירה הטבעית לפתח תאי מין מיוחדים, שהחומר הגנטי שלהם עובר חלוקת הפחתה, המשאירה בהם מערכת בודדת של מים במקום המערכת הכפולה בתאי הגוף האחרים. נוטים לחשוב שתהליך מסובך זה עבר את המסננת של הברירה הטבעית, כיוון שהוא מבטיח לצאצא 50% שונות גנטית ביחס לכל אחד מהוריו, שוני המגדיל את סיכויי הסתגלותו לתנאי סביבה משתנים ומאיץ את קצב האבולוציה. ואכן, צמחים וחיות המתרבים באמצעות רבייה מינית מייצגים את הקבוצות הגדולות והמוצלחות ביותר בטבע. אבל כיוון מחשבתה של מרגוליס לא עולה בקנה אחד עם הסברים אלה. עובדה, הבקטריות מעבירות תכונות גנטיות בין הפרטים ומשיגות גיוון גנטי באמצעות רפליקונים וסימביוזה. יתרה מזו, התהליך לפיו תא הפלואידי (מכיל מחצית המטען הגנטי) זכרי מתמזג עם תא הפלואידי נקבי (ביצית), כדי ליצור תא דיפלואידי (בעל מערכת כרומוזומים כפולה), אשר ממנו מתפתח עובר שכל תאיו דיפלואידים, לא נראה ממבט ראשון הגיוני או יעיל במיוחד.
אורך הדנ״א בכל תאי גופנו גדול פי מאה מיליון מהמרחק בינינו לירח.
על פי מרגוליס, אנו חבים את תענוגות המין לא לאסטרטגיה של גיוון גנטי, אלא לרעב. לדעתה, כל התאים האיקאריוטיים הקדומים היו הפלואידים, אבל בליעת תאים אחרים הביאה במקרים מסוימים למיזוג החומר הגנטי של הנבלע בגרעינו של הבולע, באותו האופן שמטענו הגנטי של נגיף מתמזג בדנ״א של תאינו, ועל ידי כך נעשה דיפלואידי. למצב הדיפלואידי היה יתרון בתקופות של יובש קשה, שבהן הפעילות המטבולית נמוכה, ואילו בתקופות של חיוניות סביבתית וחיפוש מזון המצב ההפלואידי היה עדיף. מכאן, סביר להניח שהברירה הטבעית עודדה התפתחותו של מנגנון מיוטי, המאפשר לתא דיפלואידי להיפטר ממטען הגנים העודף שלו בעת שהסביבה מתעוררת לחיים. אפשר עוד להניח, כי במהלך הניסויים האינסופיים של האבולוציה התקיימו זה ליד זה שני סוגי תאים איקאריוטיים: דיפלואידים בעלי מנגנון מיטוזיס, המכפיל את החומר התורשתי לפני חלוקת התא ומבטיח לכל אחד מתאי הבת את מלוא כמות הגנים, ודיפלואידים בעלי מנגנון מיוזיס, המפחית בחצי את מנת הגנים. הברירה הטבעית האירה פניה כנראה פעם לאלה ופעם לאלה, בהתאם לתנאי הסביבה המשתנים, עד אשר הופיע סוג שלישי של איקאריוטים, מצוידים בשני המנגנונים גם יחד, המסוגלים לעבור ממצב הפלואידי למצב דיפלואידי בהתאם לתנאי הסביבה ולכמות הגנים שסיפחו לעצמם מבליעת איקאריוטים אחרים. מרגוליס מאמינה שמתהליך סימביוטי זה, שבו התא הטורף לא משמיד כליל את התא הנטרף, אלא יוצר עימו מודוס ויוונדי, התפתחה המערכת המוזרה – אך היעילה – של הרבייה המינית.
תקשורת אופקית
מערכת הרבייה המינית בישרה גם את לידתה של התקשורת האנכית בטבע, שהרי תכניות המבנה של תכונות גופניות חדשות, המקודדות בחומר הגנטי, עוברות אך ורק מדור ההורים לדור הצאצאים. לעומת זאת, בעולמן של הבקטריות עובר המידע הגנטי בין פרטים מבני אותו הדור באמצעות רפליקונים. תקשורת אופקית זו עושה את כל אוכלוסיית הבקטריות לאורגניזם אחד, שכן כל תכונה גנטית חדשה הופכת תוך זמן קצר לנחלת הכלל. חילופי הרפליקונים בין הפרטים באוכלוסייה מייצגים אפוא מכניזם אבולוציוני שונה מהדארוויני, לא רק משום שהשינוי הגנטי מתפשט באוכלוסייה בכיוון אופקי, אלא גם מפני שהוא אינו נובע בהכרח ממוטציה במערך הגנים של מקבל השינוי. על ההבדל בין שני המכניזמים ניתן לעמוד מההסבר השונה שהם מספקים לתופעה שכיחה בעולם הרפואה, שאוכלוסיות חיידקים נעשות עמידות בפני תרופות אנטיביוטיות.
סוד גלוי הוא, שתרופות אנטיביוטיות 25רבות מאבדות את יעילותן במהירות יחסית עקב מוטציה גנטית מקרית באחד החיידקים. לדוגמה, די בשינוי נקודתי בגן המייצר את אחד האנזימים של החיידק כדי לאפשר לאותו אנזים לעכל את התרופה, ועל ידי כך לנטרל את השפעתה. במלים אחרות, אפילו תשמיד התרופה את כל אוכלוסיית החיידקים, הפרט שעבר שינוי גנטי ושהודות לו הוא נעשה חסין לתרופה, יוכל לייצר תוך כמה שעות אוכלוסייה חדשה של חיידקים עמידים לתרופה.
אולם מרגוליס מטילה ספק בהסבר דארוויני זה. על פי חישוביה, כדי שהאנזים הספציפי יתפתח ממוטציה מקרית, עלינו להמתין מיליון שנה. לדעתה, העמידות לתרופה האנטיביוטית מתפשטת במהירות בקרב אוכלוסיית החיידקים הודות לתקשורת האופקית של הרפליקונים. יתרה מזו, רשת תקשורת זו מפיצה מוטציות גנטיות שקורות לא רק באוכלוסיית חיידקים מקומית המזהמת מחלקה מסוימת בבית חולים, או אפילו את בית החולים כולו, כי אם גם כאלה שקורות במושבות רחוקות ושונות של חיידקים. למשל, הגן האחראי ליצירת האנזים מעכל הפניצילין יכול להימצא הרחק מבית החולים, במושבת בקטריות – שאצלן הוא משמש אולי לפירוק רקבוביות של צמחים. באמצעות רוח או בעלי חיים חודרות כמה בקטריות כאלו לבית החולים, שם הן מתערבבות עם החיידקים המקומיים ומעבירות להם את תכונת העמידות לתרופה באמצעות רפליקונים. הוכחה משכנעת לתפקיד התקשורת האופקית בפיתוח תכונות חדשות וביצירת סימביוזה בין אויבים מושבעים התגלתה במקרה לז׳ון קוונג מאוניברסיטת טנסי. באחד הימים קיבל קוונג משלוח חדש של אמבות למעבדתו. הוא הניח אותן בצלוחית נפרדת, ליד האמבות ששימשו את מחקריו, אך אחרי כמה ימים גילה שכל האמבות במעבדתו נתקפו במחלה קטלנית. בדיקה מיקרוסקופית הראתה שהוותיקות נדבקו בבקטריות שנשאו האמבות החדשות. למזלו של קוונג לא מתו כולן, אבל גם האמבות ששרדו היו חלשות מאוד, ומה שחשוב יותר – הן היו פגיעות לאנטיביוטיקה, בניגוד לעמידותן של האמבות הוותיקות. לקוונג המופתע לא נותר אלא להסיק, כי התכונה החדשה הובאה על ידי הבקטריות. כיוון שכך, אין לייחס תכונה זו למוטציה בגנים של האמבות, אלא לתהליך סימביוטי שהתרחש ביניהן לבין הבקטריות. קוונג פתח במבצע שיטתי של השגחה לריפוי האמבות הנגועות, ואמנם תוך כמה שנים הצליח לפתח מהאמבות הנגועות זן בריא, אף שכל פרטיו נשאו בגופם 40 אלף בקטריות. מה שקרה הוא, שבמרוצת הדורות מיתנו הבקטריות את תכונותיהן הקטלניות כדי להתאים עצמן לחיים המשותפים עם האמבות. התהליך הסימביוטי, מסבירה מרגוליס, הפך את הבקטריות לאברונים חיוניים במערכת האורגנית של האמבה. הצעד הבא היה מפתיע לא פחות. קוונג עקר את הגרעינים מכמה אמבות נגועות בבקטריות ושתל אותם בגופן של אמבות חדשות, נקיות מבקטריות, ואילו את גרעיניהן של אלו הוא שתל בגופן של האמבות הנגועות. והנה התברר שהאמבות הנגועות קיבלו את השתלת הגרעין ללא בעיות, בעוד האמבות הנקיות גססו ומתו ארבעה ימים אחרי מועד ההשתלה. רק אחרי שהזריק לאמבות הגוססות מנה של בקטריות הן התאוששו וחזרו לאיתנן. מסקנה: אמבות שהתנסו בחיים משותפים עם בקטריות שוב אינן יכולות לחיות בלעדיהן. הבקטריות שהיו לפנים אויב קטלני נעשו בעקבות הסימביוזה לעצמם ובשרם של צאצאי האמבות.
תופעת השתפנות מפריכה את מיתוס המאבק לחיים ולמוות שעולה מסיסמתו הידועה של ספנסר על שרידות הכשירים. סיסמה זו עיוותה כליל את כוונתו של המקורית של דארווין, שכן כשירות גבוהה אצלו לא באה לציין אלימות ותוקפנות, שבאמצעותן כביכול מפלס הפרט את דרכו במלחמת הקיום, אלא את ניצחונו בתחרות על הולדת צאצאים. בתחרות זו לא מנצחים בהכרח הגדולים, החזקים וה-תוקפנים, אלא מי שאימצו אסטרטגיות יעילות יותר. למשל, אצל מינים שונים של דגים המיושם להצלחה הוא להיות קטן וזריז. זכר צעיר, המקדים למצוא לו נקבה, נצמד לגופה וחי על חשבונה כטפיל זעיר ומצומק תמורת הזרע שהוא מעניק לה. החיים, אומרת לין מרגוליס, אינם משחק סכום-אפס, שיש בו רק מנצחים ומפסידים. המיטוכונדריות בתאי גופנו והכלורופלסטים בעלי העצים הם עדות ניצחת ליתרונו של משחק אחר, מסוג לא־סכום-אפס.
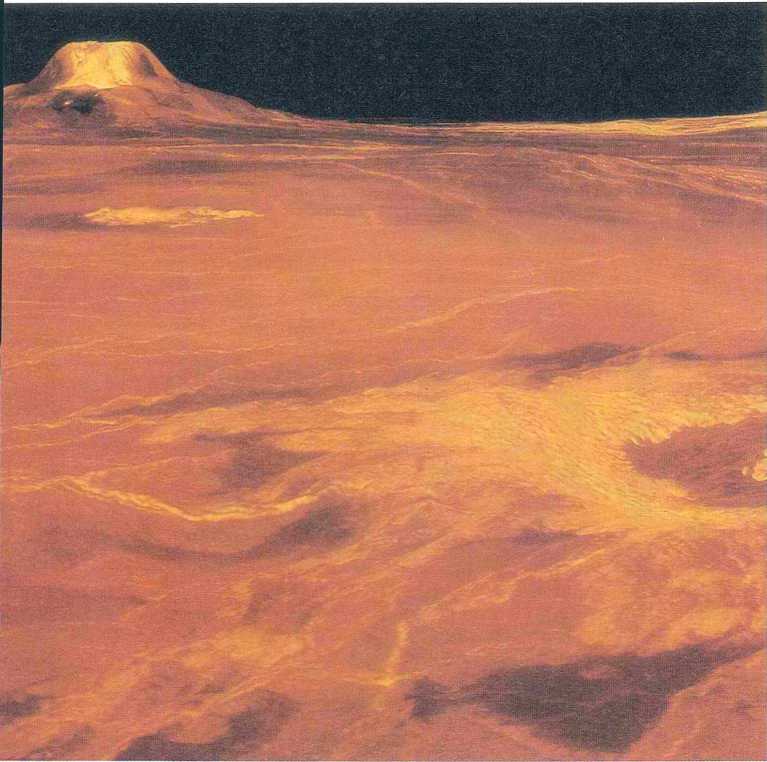
כדור הארץ כיצור חי
עולמם של החד תאים גרם ללין מרגוליס לבחון את ההיררכיה של החיים מפרספקטיבה אחרת. משנוכחה שהציפוי הדק של החיים החד תאיים לא חוסל, למרות ש-99.99% מכלל המינים שאיכלסו את כדור הארץ בשלושה מיליארד השנים האחרונות נכחדו, חברה לג׳מס לאבלוק, אבי תורת גאיה. על פי תורה זו, שהוצגה בהרחבה בשני ספריו (GAIA – a New Look at Life on Earth, 1979, ו- The Ages of GAIA, 1988), גאיה (על שמה של אלת האדמה במיתולוגיה היוונית) היא שם כולל לקליפת כדור הארץ, האטמוספירה, האוקיינוסים, הצומח והחי. כל אלה מהווים אורגניזם חי אחד, הגדול ביותר במערכת השמש. קביעה זו נשמעת בדיונית לחלוטין. עובי קליפתו החיצונית של כדור הארץ, שבקרבה מתקיימים החיים, הוא 60 ק״מ, כלומר פחות מאחוז אחד מהרדיוס שלו (6,370 ק״מ) העשוי כולו מחומר דומם. אבל, כפי שאומר הפיסיקאי ג׳רום רותשטיין, גם עץ הסקויה 26הענקי, המתנשא לגובה של יותר ממאה מטר ועוביו יותר משלושה מטרים, מורכב משכבה דקה של תאים המייצרים את קליפתו ומ-99% חומר מת, ובכל זאת אי אפשר לכפור בחיוניות החיים המרשימה של עץ מלכותי זה.
גאיה גורסת שהחיים דואגים ליציבות התנאים הכימיים, הפיסיקליים והאקלימיים החיוניים לקיומם התקין, על ידי ויסות דו תחמוצת הפחמן והחמצן באוויר ופירוק המינרלים בסלעים ובאוקיינוסים. תפיסה זו, שהחיים מעצבים באופן פעיל את סביבתם ויוצרים יחד איתה מערכת משולבת אחת, עשתה את מרגוליס בעלת ברית טבעית של לאבלוק, שכן היא ראתה בגאיה ביטוי גלובלי לרוח השתפנות שגילתה בקרב המיקרואורגניזמים.
תופעת השתפנות מפריכה את מיתוס המאבק לחיים ולמוות של האבולוציה.
עיצוב הסביבה הפיסיקלית והכימית על ידי החיים אינו עולה בקנה אחד עם הדארוויניזם, הרואה בחיים ובסביבה שתי מערכות נפרדות, המקיימות ביניהן קשר חד סטרי. לפי הדארוויניזם, הסביבה מעצבת את החיים ולא להיפך. יתרה מזו, הברירה הטבעית גוזרת את דינם של החי והצומח לחיים או להכחדה על פי מידת התאמתם לסביבה. משמע, בעלי החיים ממלאים תפקיד סביל. חיה אינה מגדלת פרווה בתגובה לקור; היא תינצל מהקור אם יש לה מראש תכונה להצמחת פרווה. הנחתם של לאבלוק, מרגוליס ועמיתיהם, שגאיה דואגת לתנאים סביבתיים מתאימים לקיום החיים, מחשידה תורה זו בכוונות ומטרות, שמקומן בכתבי דת ולא בספרי מדע. גם העובדה שכל מיני כיתות הוליסטיות ואקולוגיות אימצו את גאיה וייחסו לה איפיונים טליאולוגיים אך חיזקה את הסתייגותו של הממסד המדעי מתורה זו. אף על פי כן, אין הוא יכול להתעלם ממנה. גאיה מציגה למדע הממוסד שלוש קושיות מרכזיות: מדוע שיעור החמצן באוויר עומד על רמה יציבה של 21%; כיצד קורה שהטמפרטורה הממוצעת על פני כדור הארץ נשמרת ביציבות זה מאות מיליוני שנים סביב ˚16 צלזיוס, למרות שתפוקת החום של השמש גברה למן הופעת החיים ב-30%-25%; מדוע מליחות המים באוקיינוסים אינה עולה על 6%, שיעור שמעבר לו החיים הימיים אינם אפשריים, למרות המלחים הרבים הנשטפים בהתמדה לימים.
על פי גאיה, התשובה לשלוש השאלות הללו נעוצה במנגנוני הוויסות שמפעילים צמחים ובעלי חיים על סביבתם הכימית והפיסיקלית, כדי לשמור על התנאים האופטימליים לקיום החיים על כדור הארץ. אלמלא גאיה היו התנאים האקלימיים והאטמוספיריים בכוכב הלכת שלנו דומים מאוד לאלו של מאדים ונוגה – שכנינו במערכת השמש. הטמפרטורה על פני מאדים היא ˚53- צלזיוס, וזו שבנוגה היא ˚459 צלזיוס, ואילו הרכב הגזים באטמוספירה שלהם כמעט זהה: 95%; 96.5% דות״פ (דו תחמוצת הפחמן), 2.7%;3.5% חנקן וכמעט אפס חמצן. לעומתם, הטמפרטורה הממוצעת על פני כדור הארץ היא ˚16 צלזיוס, ואילו האטמוספירה שלו מורכבת מ־79% חנקן (לערך), 0.03% דות”פ ו-21% חמצן. אם שיעור החמצן היה גבוה מ-25%, אפילו עצים ספוגי מים היו עולים באש מלהבת גפרור, ופירוש הדבר שלא היו לנו יערות. ולהיפך, מתחת ל-15% בעלי חיים גדולים היו מתים בחנק, ויערות היו מתנוונים תחת המרבץ העבה של עשבים ועלים מתים. זאת ועוד, אפקט חממה מצטבר, שהיה נוצר מריכוז של 98% דות״פ באוויר, היה מעלה את הטמפרטורה של כדור הארץ ל-˚300 צלזיוס והופך אותו ליורה רותחת. גם אם נביא בחשבון את השפעתם המצננת של האוקיינוסים ויערות העד על האקלים, באמצעות תהליכי התאיידות ושאיבת הדות״פ מהאטמוספירה, נותרת בעינה תעלומת הטמפרטורה הנמוכה. על פי הערכתם של כמה קלימטלוגים בכירים, 27הטמפרטורה הממוצעת על פני כדור הארץ בעת הופעת החיים היתה ˚23 צלזיוס. בהתחשב בכך שתפוקת השמש עלתה מאז ב-25%, וששיעור דות״פ באטמוספירה היה גבוה עד פי 1000 מרמתו הנוכחית, הטמפרטורה הממוצעת היום היתה צריכה להיות ˚54 צלזיוס, כלומר מעבר לטווח האפשרי לקיום חיים, ולא ˚16.
של מי התרמוסטט?
כיצד אם כן מווסתים החיים את שיעור החמצן באטמוספירה של כדור הארץ ואת הטמפרטורה שלו? לטענת גאיה, המפתח טמון בשליטה על שיעור הדות״פ באוויר, שהיא המקור העיקרי של הפחמן בתהליכי החיים וגם ממלאה תפקיד מרכזי (לצד גז המתאן) ביצירת אפקט החממה. אפקט זה נובע מכך, שחלק מקרינת השמש הפוגעת בכדור הארץ וניתזת חזרה לאטמוספירה לא מתפזרת בחלל החיצון, אלא נספגת על ידי דות״פ באוויר. כתוצאה מכך נעטף כדור הארץ בשמיכה חמה, בדומה לפרחים ולירקות מתחת לתקרת הזכוכית או הפוליתילן של החממה. מכאן יוצא, שכמות הדות״פ באוויר קובעת את רמת הטמפרטורה על פני כדור הארץ: כשהיא עולה הטמפרטורה עולה, וכשהיא יורדת הטמפרטורה יורדת. השאלה היא מה גורם לה לעלות ומה גורם לה לרדת, כלומר מהו המכניזם המווסת את הטמפרטורה של כדור הארץ. שאלה זו עומדת במרכז המחלוקת בין הגיאופיסיקאים לאנשי גאיה. הראשונים טוענים לקיומה של משאבה גלובלית, המווסתת את הדות״פ באטמוספירה על פי חוקי הכימיה והפיסיקה, ואשר החיים ממלאים לגביה תפקיד שולי וזניח. שני המרכיבים העיקריים של משאבה זו הם הרי געש הפולטים דות״פ ומעלים את שיעורו באוויר, וסלעי סיליקט סידניים הבאים במגע כימי עם הדות״פ ויוצרים איתו תרכובות של סידן פחמתי. פעולת הוויסות של מנגנון פיסיקו-כימי זה עובד כך: הרי הגעש פולטים דות״פ לאטמוספירה ומגבירים בכך את אפקט החממה; עליית הטמפרטורה מאיצה את קצב ההתאיידות של מי הים, שממנה נוצרים עננים המביאים גשמים; הגשמים השוטפים את הסלעים מגבירים את קצב הריאקציה הכימית בין הדות״פ והסלעים, קושרים כמויות גדולות יותר של דות״פ בסידנים פחמתיים, ומקזזים בכך את עודף הדות״פ שנוצר מהפעילות הוולקנית; הירידה בשיעור הדות״פ באוויר מצמצמת את אפקט החממה וגורמת לירידת הטמפרטורה (ר׳ גם מכניזם הקרחונים, במאמר על ״הגיאולוגיה הדינמית של האקלים” בגיליון זה). אנשי גאיה דוחים הסבר זה בטענה, שעל פי המודל הגיאופיסי שיעור הדות״פ באוויר היה צריך להיות גבוה פי מאה מרמתו בפועל. להערכתם, ניתן ליישב את המודל שלהם עם המציאות כאשר מכניסים לתרמוסטט הגיאופיסי את החיים. לחיזוק טענתם הם מצביעים על כך שהאדמה באזורים צמחיים מכילה דות״פ בריכוז פי 40-10 מאשר באטמוספירה, כיוון שהאורגניזמים החיים פועלים כמשאבה ענקית, היונקת את הדות״פ מהאוויר וקוברת אותה דרך שורשיה בעומק האדמה, ואז הדות״פ באוויר מתמעט והטמפרטורה יורדת.
לכאורה, כל שעלינו לעשות כדי לאמת טענה זו הוא לבדוק אם עליית הטמפרטורה בתקופת זמן כלשהי מלווה בפעילות מוגברת של החיים, פעילות שאמורה לצמצם את שיעור הדות״פ באוויר ולהביא בכך לירידת הטמפרטורה. אבל הגיאופיסיקאים מצידם יכולים להסביר שעליית הטמפרטורה מאיידת כמויות גדולות יותר של מים, ואלו הופכות לגשם הגורף עימו דות״פ מהאוויר וגורם לירידת הטמפרטורה.
אצות אלטרואיסטיות
מצב העניינים הוא כזה, שהמודלים של גאיה אינם יכולים לפי שעה לספק הוכחה מכרעת לפעולתה המווסתת. גרוע מזה, מנגנוני הוויסות שלה נגועים לכאורה באלטרואיזם, ודי בחשד זה כדי לקומם נגדה את הקהילה המדעית. לדוגמה, הכל מסכימים שגשמים ושיטפונות סוחפים איתם גופרית מהיבשה ונושאים אותה לאוקיינוסים. אף על פי כן, עצים ובעלי חיים יבשתיים אינם סובלים ממחסור בגופרית. גאיה גורסת שהאיזון מושב לקדמותו באמצעות אצות מיקרוסקופיות הצפות על פני האוקיינוסים. אצות אלו מרכיבות מהגופרית במים דימתיל סולפיד (DMS), שלאחר מותן הוא מתנדף באוויר ונישא ברוח ליבשות.
הסבר זה גורם לביולוגים להרים גבה. איזה עניין, הם שואלים, יש לאצות ימיות לפתח מנגנונים המשרתים את האינטרסים של עצים, ג׳ירפות ובני אדם על היבשה? ממתי מעודדת הברירה הטבעית אלטרואיזם לשמו? ובכלל, איך יודעות האצות שבעלי החיים היבשתיים זקוקים לגופרית? מחנה גאיה משיב, כי ה-DMS משרת בראש ובראשונה את האינטרסים של האצות, שכן הוא מגן עליהן מפני התייבשות. יתרה מזו, ה-DMS שנפלט לאטמוספירה גורם להתעננות (בדומה ליודיד הכסף, המשמש אותנו להורדת גשמים), וזו מעוררת רוחות עזות המערבלות את שכבת המים העליונה של האוקיינוסים ומעשירות את מצע המזון של האצות. הסבר זה מספק אפוא לברירה הטבעית שתי סיבות טובות לעודד את מנגנון ה-DMS באצות.
מנגנון ה-DMS לא הוכח, ואף יש ממצאים המורים על כך שפליטת ה-DMS לאטמוספירה אינה פרופורציונלית לצפיפות האצות באוקיינוס. אך הבעיה המרכזית של לאבלוק ומרגוליס נעוצה בניסיונם לייחס לגאיה פעילות שיטתית ליצירת תנאי סביבה מיטביים לחיים, ניסיון שבמקרים רבים אינו תואם את המציאות, שכן ידוע כי מינים שונים הורסים בשיטתיות את סביבתם ומסכנים בכך את קיומם. זאת ועוד, התנאים המיטביים למינים השונים אינם זהים, ומאחר שאיש אינו מעלה בדעתו שגאיה מעדיפה מין אחד על משנהו, נשאלת השאלה על פי מה נקבעו ערכי החמצן והטמפרטורה. יש מינים המשגשגים ב10- צלזיוס, ויש הקופאים בו למוות; יש אורגניזמים החיים היטב באוויר המכיל 40% חמצן, ואחרים – עכברים, למשל – שורדים היטב באוויר רווי חד תחמוצת הפחמן. יתרה מזו, אם גאיה פועלת למען בריאותו הכללית של כדור הארץ, מדוע עודדה את הופעת הסיאנובקטריות, שהרעילו את האטמוספירה בחמצן והביאו בכך שואה על רוב החד תאים באותה עת? אם התשובה היא שבטווח הארוך יש לחמצן פוטנציאל חיובי, כי הוא מאפשר לבעלי חיים גדולים 28להתפתח, משמע שגאיה לא רק מחוננת בכושר חיזוי, כי אם גם נושאת באמתחתה מטרות ותכליות. הוא הדין בטענה שרמת החמצן הנוכחית באוויר מיטיבה עם היערות, כי היא מאפשרת פריצתן של דליקות מבוקרות, השורפות את מרבצי העלים ומחסלות עצים זקנים. אנשי גאיה טוענים, שמינים שונים של עצים משקיעים 5% מהאנרגיה שלהם ביצירת חומר דליק ביותר (איזופרן), המסייע להתפשטות האש. מכאן משתמעת מסקנה בעייתית, שהעץ האינדיווידואלי מסכן את קיומו כדי לקדם את האינטרס הכללי של היער. סוג זה של הסברים מחשיד את אנשי גאיה בפנגלוסיזם (על שמו של ד״ר פנג-לוס, מורו ורבו של קנדיד): עולמנו הוא הטוב שבעולמות, שהרי האורגניזמים החיים עימנו עמדו בהצלחה במבחן הברירה הטבעית, ועל כן הם מייצגים את הפתרון הטוב מבין כל הפתרונות האפשריים. הדארוויניזם דוחה כמובן פנגלוסיזם זה ומציג גירסה משלו: עולמנו הוא אכן הטוב שבעולמות, אבל רק למי שהסתגלו אליו. אילולא היה הדבר כך, היינו נאלצים למשל להצביע על האנטומיה של העכבישים כעל תכנית המבנה הטובה מבין כל התכניות האבולוציוניות האפשריות, שהלא העכבישים חיים עימנו עד עצם היום הזה. הבעיה היא שהעכבישים לא יכולים להגדיל את מוחם מעבר לנפחו הנוכחי, משום שהוא מוקף בוושט שלהם, ומצד שני – הם ניזונים אך ורק מנוזלים, מאחר שמוחם לוחץ על הוושט. קשה להצביע על מילכוד זה כעל הפתרון הטוב מבין כל הפתרונות האפשריים.
החיים אינם משחק סכום־אפס, שיש בו רק מנצחים ומפסידים.
לזכותו של לאבלוק ייאמר, שהוא לא מתיימר לתת תשובות לכל השאלות. בכנות ראויה לשבח הוא הודה לא אחת שניסוחיו הכוללניים, המתארים את כדור הארץ כאורגניזם הפועל למען החיים, קרובים יותר לפיוט מאשר לאמירה מדעית מוצקה. יתרה מזו, לאבלוק מעיד על עצמו שהשאלה אם תורת גאיה נכונה או שגויה היא בעיניו בעלת חשיבות משנית. החשוב הוא שהיא מציגה התייחסות חדשה ופורייה יותר כלפי כדור הארץ וכוכבי לכת אחרים, ומביאה את הביולוגים, הגיאולוגים והגיאוכימאים להתמודד עם שאלות ועם תופעות שלא נתנו דעתם עליהן. ואמנם, גם מבקריה החריפים של תורה זו מודים, כי היא חוללה זרם של גירויים אינטלקטואליים בכמה דיסציפלינות מדעיות. ואולי חשיבותה נעוצה במסר המרכזי שלה, שהחיים והסביבה מקיימים שיתוף פעולה הרבה יותר עמוק מכפי שאנו נוטים לחשוב, ושהמין האנושי אינו הבעלים של כדור הארץ, אלא רק אחד מדייריו. בתור שכזה חובה עליו לשמור על רשות הרבים כאילו היה רשות הפרט, באשר השניים אינם אלא אחד. ■ צ. י.

