





הקדמה:
פעילותו של האדם בטבע, פעילות הנעשית לעיתים ללא תיכנון מספיק וללא מחשבה מסודרת, מעוררת אצל רבים מאיתנו שאלות הקשורות במשמעות של האירגון בטבע, בפשר ההתערבות של האדם במבנהו ובאפשרות לחזות את עתידו. האם הטבע חייב להיות מסודר, מאוזן וחזוי מראש? האם אי סדר הוא גורם שלילי בטבע או שמא יש לו תפקיד חיובי בו? הייתכנו חיים במצב של אי ודאות? האם לתהליכים סביבתיים מקריים ובלתי חזויים יש תפקיד בעיצוב עולם החי? ומעל לכל, הייתכן שמערכת כל כך מאורגנת, כמו אורגניזם חי, עשויה להיות בו בזמן גם חזויה מראש וגם כאוטית?



ברצוני להשתמש ב״זכות הספק״ ולהעלות הירהורים אחדים בדבר הרלוונטיות של השימוש הגובר במושגים כמו ״חוסר ודאות״ ו״כאוס״ במערכות ביולוגיות, ובדבר המגבלות של התיאור הפרקטלי שלהן.
בשנים האחרונות הפך העיסוק ב״כאוס״ לאופנתי, לא רק במתמטיקה ובפיזיקה אלא גם בביולוגיה. אולם אני מטיל ספק אם למושג כמו כאוס יש משמעות בביולוגיה ובהבנת החיים.
כדי לקיים את החיים צריך כושר חיזוי, אבל כדי שאפשר יהיה לחזות מראש מצב דרוש סדר, דהיינו חוקיות של אירועים, וכדי לקיים סדר – צריך מידע. סדר הוא מצב של מערכת, המתאפיין ברמת אנתרופיה נמוכה1 , שקיימים בו קשרי גומלין בין המרכיבים השונים של המערכת, ועל בסיס המידע הזמין להם מתארגנים מרכיבים אלה בצורה קבועה וחוזרת.
אילו היו המולקולות של החומר החי מגיבות זו עם זו בצודה מקרית, צפוי היה שייווצר מספר אינסופי של צירופים. אבל אין זה כך. עובדה היא, שמספר הצירופים הבין מולקולריים הקיימים בחומר חי הוא קטן למדי. תכונות ביוכימיות רבות, כמו ההרכב הבסיסי של החלבונים או מרכיבי חומצות הגרעין, הן אפילו זהות לכל האורגניזמים החיים המוכרים. לדוגמה, החלבונים של מרבית האורגניזמים החיים עשויים מחומצות אמינו ״שמאליות״ (L-amino acids) ולא מחומצות ״ימניות״.
אני מטיל ספק אם למושג כמו כאוס יש משמעות בביולוגיה ובהבנת החיים
לעומת זאת, חומצות הגרעין מורכבות דווקא מסוכרים ״ימניים״ (D-sugars) ולא מ״שמאליים״. הדמיון בתכונות אלה במערכות חיות, ואף ביטויים אתרים של סדר בהן, לא יכול היה להיווצר במערכת כאוטית, שבה ״אין שני מצבים שווים״.
דוגמה נוספת לאחידות ולקביעות של צורות היסוד של החיים היא צורת הסליל.
סליליות בצמחים מתבטאת בגדילי הדנ״א, בהשקעת אגדי מולקולות התאית (פיברילות) בדופן התא, בסידור תאי הגבעול, בסידור העלים לאורך הציר, בסידור הפרחים בתפרחת, באתרי יציאת השורשים הצדדיים בשורשים ועוד. סליליות פירושה חוקיות. לפיכך, הכרת מקומם של שלושת הפרחים הראשונים בתפרחת של חמנית והכרת החוקיות של הסליל בצמח זה מאפשרת חיזוי מדויק של מקומם של שאר הפרחים. זית הוא זית, משום שהצורה של עליו ועמדתם הן קבועות ואינן אקראיות, משום שאירגון העלים על הגבעול הוא נגדי, משום שלפירותיו צורה מסוימת, טעם מסוים ותכולת שמן מסוימת, ומשום שאבקתו מאופיינת דווקא על ידי חלבונים אלרגניים (גורמי אלרגיה) מטיפוס מסוים. הסדר העוקב שבו נוצרים עלי הזית ניתן לחיזוי, בתנאי שידועים לנו הנתונים הסביבתיים כמו טמפרטורה, עוצמת ההארה, משך היום, זמינות האנרגיה המטבולית או משך ההתפתחות. החוקיות ביצירת איברי צמח, שמשמעותה הוא העדר של גורמים כאוטיים, היא אפוא ברורה וניתנת לחיזוי. דגמי היסוד חוזרים על עצמם.

ראשית החיים: יצירת סדר מכאוס
אין ספק שהחיים נוצרו מתוך סביבה כאוטית. אבל יש להבדיל בין המצב הכאוטי ששרר בזמן היווצרות החיים ובין כל שאר המצבים ששרדו לאחר מכן. ההנחה המקובלת כיום היא, שלפני כארבעה מיליארדי שנים אירע על פני כדור הארץ המתהווה, הכאוטי, תהליך שהביא בעקבותיו ליצירת הסדר הראשוני של החיים.
העובדה שלפני היווצרות החיים שרר מצב כאוטי אינה רלוונטית להסבר אירגונם או תיפקודם של החיים כפי שהם היום. אמנם, יש המשערים שחיים נוצרו יותר מפעם אחת, אבל הדמיון המפליא במבנה ובהרכב של כל צורות היסוד של החיים המצויות היום, בדרך פעולתם של אלה ובכיוונם מעמיד בספק את תקפותה של השערה זו.
החיים הם אחד המרכיבים הבסיסיים של הטבע, ועם זאת הם שומרים על ייחודם בתוכו. אורגניזמים חיים אינם מתאימים את עצמם למרכיבים הדוממים שבטבע. להיפך, החיים הם ביטוי ליכולתו של התא החי שלא להתאים את עצמו לסביבה. זאת היכולת שלהם להסתגר בתוך ״גטו״ מאחורי הממברנות הבררניות ולקיים את עצמם בסביבה פנימית קבועה, שהם עצמם מייצרים אותה. היכולת ליצור סביבה פנימית מסודרת וקבועה (הומיאוסטזיס) היא אשר הגנה על החי הראשוני מפני תנאי הסביבה ששרדו בזמן היווצרו, ואיפשרה את הישרדותו ואת המשך קיומו. ההומיאוסטזיס הוא תנאי בסיסי לקיומם של החיים.
ומה יקרה אם ייפגם הסדר ובאורגניזם יתפתח מצב כאוטי? במקדה כזה, הוא לא יוכל לבנות לעצמו סביבה פנימית קבועה ויציבה, ולא יהיה לו סיכוי להתקיים. ברגע שתהליכי החיים מתחילים לפעול בצורה שאינה ניתנת לחיזוי, חל שיבוש בתיפקוד של מרכיבי התא, בתיאום שביניהם או במיקומם. זהו הרגע שבו נגמרים החיים ומתחיל המוות.
הצורך בחיזוי אינו כל כך בעייתו של הביולוג החוקר את המערכת מבחוץ; הוא בעיית הקיום של המערכת עצמה. כל אחד מאלפי המרכיבים של המערכת החיה חייב לחזות מראש את מקום פעולתם של האתרים, את שיעורי פעולתם ואת תוצרי פעולתם, ולתאם את פעילותו בזמן אמיתי. התא החי חייב להיות מסודר ומאורגן בצורה מוחלטת כדי שיתפקד ויחיה. יש להדגיש, כי כשמדובר בסדר וביכולת חיזוי, אין הכוונה למצב סטטי חסר גמישות, אלא במבנים ובתהליכים דינמיים, המתקיימים באורגניזמים חיים.
כל מערכת תיפעולית צריכה לדעת מתי להגיב למידע ומתי להתעלם ממנו. עליה להיות גמישה בתגובה לשינויים סביבתיים, אך בו בזמן היא חייבת לשמור על הנוקשות של תבניות התנהגות קבועות וחזויות. לכן, צבירת מידע ובוודאי העברתו מבטאים סוג מסוים של סדר, החיוני לקיום החיים. מידע הוא חסר משמעות, אם האותות שלו נעדרי אירגון. מזה נובע, שמערכות כאוטיות, כלומר מערכות שאינן חוזרות על עצמן ושלא ניתן להגדיר את החוקיות שלהן, אינן מסוגלות לקלוט מידע מסביבתן. כלומר, אם מערכת כזאת ״לומדת״ משהו ומתנהגת בהתאם, היא מפסיקה להיות כאוטית והופכת להיות ״מערכת חזויה״. מאחר שחיים פירושם התארגנות בסביבה פנימית קבועה, קיים ניגוד בסיסי בין ״חיים״ ובין המצבים של אי ודאות ושל חוסר היכולת ״ללמוד״, שמייחסים למערכות כאוטיות.

החיים הם תכונת יסוד, ועצם קיומם מותנה בתיפקודן של מערכות משוב חזויות (ליבוביץ, 1994; זיו, 1993). לכן, גם אם הסדר הראשוני של החיים נוצר במהלך כאוטי כלשהו -בכך הסתיים תפקיד הכאוס בביולוגיה. מידע, בדיוק כמו סדר ואירגון, דורשים השקעת אנרגיה, אבל ברגע שנרכש המידע הראשוני, השקעת האנרגיה הדרושה לפיתוח המידע ולשימוש בו קטנה לאין שיעור.

מטבוליזם, גנטיקה ואנטי כאוס
אורגניזם חי אינו סובל מצבי אי ודאות. התהליכים הבסיסיים ביותר שלו מתאפשרים בזכות אירגונם של 37מרכיביו: מים, סוכרים, מינרלי מזון, מולקולות co2 וכיוצא באלה במערכות מסודרות. כבר צורות החיים הראשונות נאלצו להיבדל מתוך התמיסה המימית החופשית שזרמה סביבם, בגלל האי ודאות ששרדה בסביבה זו. הפתרון שנמצא למערכת האירגון הראשית היה כנראה בהיצמדות לשטח פנים אחיד, יציב ומסודר. יש השערה, שהיתה זו תבנית העשויה ממינרלי חרסית. הפרדת הסביבה הפנימית שלהם מהסביבה החיצונית, הכאוטית, נעשתה באמצעות ממברנות בררניות.
הסדר דרוש לא רק לשם קיומו של אורגניזם חי יחיד, אלא גם לקיומו של הטבע כולו
גורם נוסף ביצירת האפשרות לקיום חיים, אשר גם הוא אמור להיות מסודר וחזוי, הוא היכולת הקטליטית שהם פיתחו – היכולת לזרז תגובות ביוכימיות בתנאים פיזיולוגיים מתונים. קצב התהליכים הכימיים היה איטי מדי לצורך קיום חיים, והיה הכרח לזרז אותם, תוך שמירה על סביבה פנימית מוגדרת ויציבה.
פתרון זה היה כרוך ביצירת אנזימים – חלבונים בעלי אתרי פעילות שלהם מבנה מדויק מאוד – שבלעדיהם לא היו חלים התהליכים המטבוליים בטמפרטורה פיזיולוגית. ברור שסביבה כאוטית הורסת את הסיכוי לפעילות אנזימטית מבוקרת.
אלה הן דוגמאות לפעילויות אנטי כאוטיות של המערכות החיות, שגורמות להורדת האנתרופיה שלהן. פעילות אנטי כאוטית כזאת היא האמצעי שאיפשר גם את קיומם של החיים וגם את האבולוציה שלהם.
דוגמה נוספת לפעילות אנטי כאוטית ניתן לראות בתיפקודה של המערכת הגנטית. כל תא חי מכיל מכלול של גנים. אולם, אילו כל הגנים היו פעילים בו זמנית, היה נוצר אי סדר בתא, ובעקבותיו מוות מיידי. לכן קיים באורגניזם החי ויסות של פעילות הגנים השונים, ולמעשה רק אחדים מהם פעילים כל הזמן.
חוקיות ברורה קיימת ב״הדלקה״ או ב״כיבוי״ של גנים, וכל סטייה מחוקיות זו מדוכאת על ידי המערכת עצמה.
השקעה במניעת טעויות ושינויים עדיפה מהשקעה בתיקון. ובאמת, מניעת שינויים בטמפרטורה, החיונית כל כך לבעלי דם חם, או הגנה מפני עודפי רדיקלים חופשיים, הנעשית בתאים המטמיעים של הצמח, הן קו ההגנה הראשון בשמירת הסדר באורגניזם חי.
אבל אם מנגנונים אלה אינם מספיקים ונגרם שיבוש בפעילות, נכנסים מנגנוני התיקון לפעולה. סדירות פעילותה של מערכת המידע הגנטית מותנית לא רק במניעת שינוי משמעותי, אלא גם בטיפוח האפשרות לתיקון סטיות. ואכן, בתא החי התפתחו מערכות אחדות של תיקון. נזכיר כאן מביניהן למשל את מערכת התיקון הכללית, הפועלת על כל הרצף של ה-DNA. תפקידה של מערכת זו הוא לזהות מהר ככל האפשר את המקום שבו נגרם נזק, להגיע אליו ולהרחיק את הבסיס השגוי מתוך הרצף. מערכת התיקון הזאת בנויה מחלבונים ״חכמים״, המכניסים את היחידה הפגומה לטבעת, והיא מפורקת אחר כך יחד עם הפגם. הקטע ה״בריא״ של ה-DNA נתפר אז מחדש.
גם תיקון פגמים במערכת הגנטית של אורגניזם חי אינו מקרי. התיקון הוא מתוכנן, והוא מתבסס על כך שה-DNA הוא דו גדילי. כל תיקון דורש השוואה למקור. זוג גדילי ה-DNA הם אפוא לא רק התבנית הקובעת את המידע הגנטי, אלא גם מערך, אשר במקרה של קילקול של גדיל אחד, שומר את הגדיל השני לצורך ״גיבוי״ ותיקון נכון. עם זאת, גם החייט הטוב ביותר לא יתקן חליפה מבלי שניתן יהיה לזהות את התיקון. גם בתא החי, התיקון במערכת הגנטית אינו תמיד זהה בכל מאת האחוזים למקור, ולעיתים יש מידה מסוימת של סטייה. אולם רק סטייה שאין בה כדי להפריע להמשך קיומם החזוי של תהליכי החיים, היא שתשרוד.
כיצד התארגנה הפעילות האנטי כאוטית בפעם הראשונה, בצודה מקרית או בצודה דטרמיניסטית? זאת אין אנו יודעים, אבל מפתה להניח, שהיה זה אחד מגירויי הסביבה המסודרים, שנתן את המענה לצורך ראשוני זה באירגון ובמידע. גירוי כזה יכול היה להינתן על ידי הכיוון של שדה הכבידה, על ידי שינויים קבועים של עונה (יום ארוך וטמפרטורה גבוהה של קיץ המוחלפים בחוקיות ברורה ביום קצר וטמפרטורה נמוכה של חורף), על ידי המקצב החוזר של הגאות והשפל (שינויים יומיים, דו-שבועיים או שנתיים), על ידי חשיפה לרצפים של גלים המתנפצים על החוף וכדומה. אמנם גירויים אלה יכלו להעביר מידע מועט בלבד, אבל אורגניזם פרימיטיבי זקוק רק למעט מידע, ועשוי היה להתארגן על סמך פירורים ראשוניים כאלה. ככל שהמערכת החיה נעשתה מפותחת יותר ומסודרת יותר, כך היא דרשה יותר מידע, וההסתברות שהמידע שאצור בה נוצר באקראי קטנה והולכת.
גם הטבע הדומם אינו כאוטי
חוסר האפשרות לשרוד במצב כאוטי אינו מאפיין רק חומר חי. גם רוב המרכיבים הדוממים בטבע כפופים לחוקיות ברורה. לחוקיות קבועה חייבת להיות השפעה 38קבועה, שתוצאותיה קבועות מראש, ולכן גם השפעתה על הטבע הדומם חזויה.
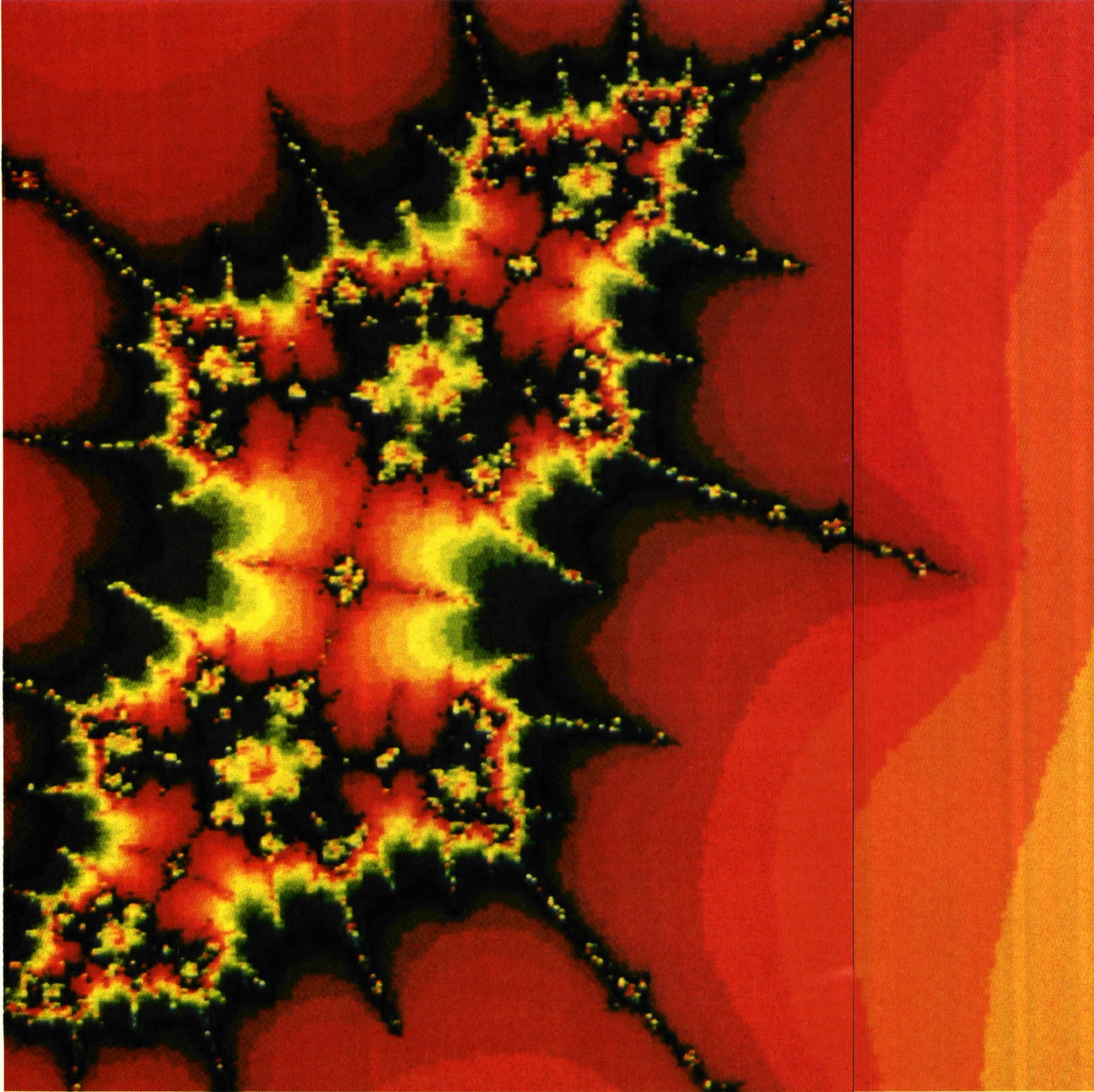
האופי הדטרמיניסטי של השפעות הסביבה, על חומר דומם, מודגם בדרך התפחותם של גבישים. מלח יכול להתגבש בצורות שונות. בסביבה לא מבוקרת נוצרים גבישים הניתנים לתיאור פרקטלי, בעוד בסביבה מבוקרת ייווצרו גבישים בעלי תכונות אחרות (יששכרי, 1994). אם הביטוי הצורתי של הגביש נוצר רק בסביבה מסוימת בעוד בסביבה אחרת צפוי שתהיה לו צורה שונה, חייב להיות בתהליך יצירתו של כל גביש של מלח מרכיב סיבתי חזוי – דטרמיניסטי, וכלל אין מדובר בתהליך כאוטי.
לפיכך, הסדר דרוש לא רק לשם קיומו של אורגניזם חי יחיד, אלא גם לקיומו של הטבע כולו. שיבוש הסדר שבו מאורגנים המרכיבים החיים שבטבע וביטול אפשרות החיזוי שלהם יגרמו לתוהו ובוהו ולמוות. מאתר שהטבע שעל פני כדור הארץ כולל בתוכו מרכיבים פיזיקליים וכימיים ומרכיבים ביולוגיים, הוא אינו יכול להישאר ״טבע כאוטי״, אם אחד ממרכיביו נפגע ומת!
ואמנם, כאשר מגיעים לניתוח אופיים של התהליכים בטבע, מתעוררים ספקות גם אצל המטיפים האדוקים ביותר של תורת הכאוס. הדבר בולט פה ושם בספרות (יששכרי, 1994; גליק, 1991). הצהרה כמו: ״כאוס מצביע על מערכות ששורר בהן משוב בלתי ליניארי״ בנויה על סתירה פנימית. אם אנו קובעים אפריורי שבתופעה כאוטית אין שני מצבים זהים, הרי לפי תורת הכאוס עצמה לא ייתכן שיימצאו בה מנגנוני משוב, שכן מנגנוני משוב מעצם טבעם כרוכים בלמידה. כאוס הקשור בלמידה אינו כאוס, אלא מערכת הניתנת לחיזוי. ייתכן שההצהרה הזאת הולמת מערכות מתמטיות מסוימות, אבל כאשר מגדירים מערכות חיות כמערכות ש״אין בהן שני מצבים זהים״, כמערכות ש״למשוואותיהן אין שני פתרונות זהים״, או כ״כאלה שהן חסרות דפוס מחזורי של תוצאות״, ברור שאין אפשרות ליישם מושג זה במערכות חיות.

דוגמה נוספת לסתירה פנימית נוכל לראות בהצהרה: ״הפיזיולוגים מצאו סדר מפתיע בכאוס המתפתח בלב האדם, שהיה הסיבה הראשית למוות פתאומי שאין לו 39הסבר״. משפט כזה רק מדגיש את הספקות שיש אפילו למחבר הספר ״כאוס״ ביחס למהות האמיתית של הסיבה. אם הסיבה למוות היא כאוטית, לא ייתכן בה ״סדר מפתיע״, ואם היא פרי תהליך דטדמיניסטי-סיבתי, הרי גם לא יכול להיות בה אלמנט כאוטי והיא גם אינה יכולה להיות בבחינת ״שאין לו הסבר״. לכל היותר ניתן לטעון במקרה זה, שההסבר אינו ידוע. גם הרצון להבין את משמעותו של הכאוס כולל פרדוקס, שכן: ״להבין את הכאוס פירושו למצוא בו חוקיות וסדר. אבל אם יש בכאוס ״חוקיות וסדר״ (דטרמיניזם), הוא חדל בכך להיות כאוס… ״ (ינאי, 1994).
השונות מגדילה את סיכויי ההישרדות של חלק מאוכלוסיית המין
גם דוגמאות רבות של תופעות ״כאוטיות״ המובאות בספרות אינן תמיד נכונות: האירגון המרחבי של העלעלים בעלה של שרך, המובא כ״דוגמה לתופעה טבעית כאוטית, היוצרת מבנה פרקטלי״, הוא דווקא דוגמה מצוינת למערכת דטדמיניסטית. הסיבה לכך טמונה בחוקיות הברורה שברצף התהליכים המבטאים את סדר התפתחותו של עלה זה. התא התחילי, הקודקודי, של אותו שרך מקבל גירוי לחלוקה במערכת פיגמנט-הורמון קבועה. מישור החלוקה של תא זה קבוע מראש, ומידע זה מתבטא הן במיקומו של גרעין התא ובעמדתו, הן בכיוון יצירתם של סיבי הכישור והן במקום שבו נוצרת לוחית התא (דופן משותפת לשני תאים לאתר חלוקת התא). בזכות הסדר המופתי של תהליך החלוקה נגזרים מתא האם תאי בת. כיוון צמיחתם של תאים אלה הוא כלפי מטה, בזוויות קבועות ובכיוונים הקבועים מראש. בתנאי סביבה מוגדרים יהיה קצב חלוקתם של תאים אלה קבוע.
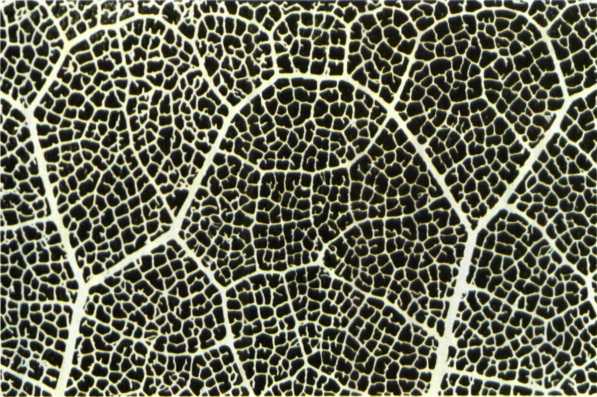
במהלך חלוקתם יוצרים התאים התחיליים רקמות מוגדרות וקבועות, שגם תכונותיהן וגם תוצריהן ניתנים לחיזוי מדויק, וכך גם התפתחות הרקמות וצורתן המוגמרת. התפתחות העלה בשרך אינה אפוא תהליך כאוטי, אלא תהליך דטרמיניסטי מובהק, המתבסס על דגם פעילות ליניארי או לוגריתמי, החוזר על עצמו. לדוגמה זאת עוד אשוב בהמשך. ועוד דוגמה: היום אנחנו יודעים, שקביעת המבנה הכאוטי כביכול של צורת העלה של העגבנייה נעשית על ידי גן אחד (זקס, 1994). שינוי במרכיב אחד של המערכת הדטרמיניסטית הזאת עשוי לתת לנו, בהתאם לרצוננו, עגבנייה בעלת עלים תמימים או מסולסלים. אילו התפתחותו של עלה כזה אכן היתה כאוטית, הוא היה הופך למסת תאים בלתי ממוינת. אבל, כפי שכולנו מכירים, צמחי עגבנייה אינם מייצרים רקמה בלתי מוגדרת (קאלוס) בראש הנצר, אלא מייצרים עלים או פירות בעלי צורה מוכרת, קבועה וניתנת לחיזוי.
יכולת חיזוי קיימת לא רק במערכות חיים ״נורמליות״ אלא גם במערכות הנראות לכאורה ״בלתי נורמליות״. התפתחותו של גידול סרטני באורגניזם חי נחשבת לשבירה של המערכת הדטרמיניסטית שלו. לכאורה, שבירה כזאת אינה מאפשרת חיזוי של אפשרות הגידול, של כיוון חלוקות התאים, של קצב גידולם או של מצבם הסופי. לכן נחשב הגידול הסרטני לתופעה כאוטית. אבל לא כך הוא המצב.
היום אנחנו יודעים, שהביטוי הצורתי (הפנוטיפי) של תאים סרטניים נקבע על ידי המרכיב הגנטי של התא, במשולב עם מרכיב סביבתי חשוב – הסביבה שתאים אלה גדלו בה קודם לכן. לדוגמה, התפתחותו של גידול סרטני בעכברים תהיה שונה אם הוזרקו להם תאים סרטניים מתרבית רקמה שגודלה במבחנה, או שהוזרקו להם תאים מרקמה זהה, אך כאלה שהתפתחו קודם לכן בגופו של עכבר אחד. נראה אפוא, שאפילו הסיכוי להתפתחותו של סרטן ניתנת לחיזוי, על סמך מקורם של התאים ועל סמך ידע מוקדם של תנאי גידולם (ויץ, 1994). למרות שהמעבר מדרגת ממאירות נמוכה לדרגת ממאירות גבוהה יכול לחול במספר רב מאוד של מסלולים, הגידול הסרטני אינו כאוטי. זהו גידול דטרמיניסטי בעל חוקיות משלו, אם כי חוקיות השונה מזו של הרקמות האחרות של האורגניזם.
דמיון, שונות, סטטיסטיקה ודיוק
הוויכוח על השאלה אם תהליך מסוים הוא מסודר ותוצאותיו ניתנות לחיזוי, או אם הוא תהליך כאוטי, מתמקד לעיתים במיקומו של הגבול המדויק שאנחנו מותחים בין סדר (כאשר השונות היא מזערית וודאות החיזוי מרבית) לאי סדר (כאשר השונות היא מרבית וכמוה גם האי ודאות). על כן יהיו מי שיאמרו: נכון שלכל אחד מהתהליכים בביולוגיה יש חוקיות וכיוון, ונכון שהממוצע ניתן לחיזוי, אבל מה עושים עם סטיות מחיזוי ״מדויק״ כזה? ואכן, אחד הדברים הפחות מדויקים במערכת חיה הוא המושג ״מדויק״. תשובה לטיעון זה חשובה לדיוננו, כי הגדרתו של תהליך כ״אקראי״ או כ״דטרמיניסטי״ עשויה להיות מותנית בדיוק החיזוי.
ככלל, מידת החיזוי שבתהליך פיזיקלי מאקרוסקופי היא 40בממוצע גבוהה, ובוודאי גבוהה מזו שבתהליכים ביולוגיים. כאשר ידוע מצב המוצא של תהליך וידועים תנאי הסביבה שלו, ניתן לחזות בסבירות גבוהה מאוד את התוצאה. ההערכה של הדיוק במערכת חיה היא שונה, והשונות של התאים היא שמספקת לה מרחב פעולה: מה שנראה לנו ״שונה״ לגבי מיקום העלעל על עלה השרך, עשוי להיות ״מדויק״ ו״מסודר״ לגבי התאים של צמח זה. אחד מגורמי השונות של המערכת החיה טבוע באופיה. לכן, לפני שבודקים את אופי התהליכים החלים באוכלוסייה חיה, צריך לוודא שאנחנו אכן עוסקים באוכלוסייה מוגדרת ושמרכיביה של אוכלוסייה זו אינם נבדלים זה מזה בצורה בלתי סבירה. אין ספק שצידו הצפוני של עץ הוא חלק מאותה מערכת תאים אשר אליה משתייך צידו הדרומי: במקרה כזה ניתן לחזות את התפתחות הענפים בצידו האחד של העץ לפי התפתחותם בצידו השני. אבל, אם התנאים הסביבתיים והחוקיות של ההתפתחות שבצידו הצפוני של העץ שונים מאלה שבצידו הדרומי, לא לניתן להשתמש באותם הכללים לחיזוי יצירת הענפים בחלקיו השונים, מכיוון שאז הם יהיו שני ״עצים״ שונים. אין מדובר כאן בכאוס אלא בשני דגמי חוקיות, מותני סביבה, שכל אחד מהם ניתן לחיזוי.

אותו הדבר חל גם בהתפתחותם של עלעלים בשרך. קיומו של המידע הגנטי הדרוש ליצירת עלעלים במקומות שונים אינו מספיק. מידע זה צריך להגיע לידי ביטוי. אם נוכל ליצור סביבה לתחיליות העלים, סביבה אוטופית, שתהיה אחידה בצורה מוחלטת, נוכל גם לחזות בדיוק מלא את מספר העלעלים שייווצרו על כל עלה ואת מיקומם. חיזוי כזה, שיתבסס על תכונות התאים התחיליים, יהיה תקף רק באותו צירוף של תנאי סביבה אחידים, ודיוקו יבטא אז את הפוטנציאל הגנטי של הצמח. התבטאות מסודרת של תופעה במערכת חיה דורשת הן את הכרת המידע האחראי לה (הגנוטיפ) והן את הכרת הביטוי הצורני-תיפקודי שלה (הפנוטיפ), התלוי בסביבה. לשינויים במידע הגנטי יש להתייחס משתי נקודות מבט: יש לראות בהם את עצם השינוי שנגרם במערכת המידע, באירגון ה-DNA, אך גם ביכולת שלהם עצמם לגרום לביטוי הפנוטיפי. אורגניזם לא ״ירגיש״ בשינוי בגנום שלו (מוטציה) עד ששינוי זה יגיע לידי ביטוי. להבחנה כזאת יש חשיבות לדיוננו, משום הטענה שיש שינויים ב-DNA, העשויים להיות מקריים. אולם אפילו במקרה כזה, הביטוי הפנוטיפי של השינויים הללו אינו מקרי כלל ועיקר. אם הכל כל כך חזוי וכל כך קבוע וכל כך מאורגן, כיצד קרה שיצורים חיים אינם זהים זה לזה, ושלמרות שכל גזר דומה לרעהו במהותו הבסיסית, עדיין הוא שונה ממנו בפרטים? ובכן, לא כל הפרטים ממין אחד הם זהים, ושונות היא תכונה בסיסית של המינים. ניתן לראות בשונות תכונה שיש לה השלכה ישירה על היכולת של אורגניזמים חיים לשרוד בתנאים המשתנים של הסביבה. אלמלא השונות, היה כל שינוי בתנאי הסביבה יכול לגרום להכחדת המין כולו; השונות מגדילה את סיכויי ההישרדות של חלק מאוכלוסיית המין. עם זאת, יש לזכור שהשונות אינה תוצאה של אי יכולתו של אורגניזם חי לייצר צאצאים זהים בסביבה זהה, אלא דווקא תכונה הסתגלותית (אדפטיבית), שכל אורגניזם מטפח אותה. שונות כזאת אינה בהכרח פרי תהליכים כאוטיים או תוצאה של תגובות אקראיות, אלא ביטוי של הפוטנציאל הגנטי הקיים בתנאי סביבה שונים. בטווח מסוים של שונות ניתן בהחלט לנבא את צורתם של האיברים השונים שיתפתחו על הצמח ואת ממדיהם. הבסיס לשונות הביולוגית טמון אפוא הן בשינויים הסביבתיים הקיימים סביב כל תא וכל פרט, והן בשונות הפנימית, ושני המרכיבים תורמים בסופו של דבר ליצירת סדר.

דטרמיניזם, כאוס ואבולוציה
אירגון החיים לא נותר כמות שהוא בארבעת מיליארדי השנים שעברו מאז היווצרם. אילו היה האירגון דטרמיניסטי בצורה מוחלטת והחיים היו נותרים ללא התפתחות, היינו היום כולנו מעין ארכיבקטריות. כיוון שאיננו כאלה, ברור שקרתה התפתחות – אבולוציה. זהו השטח שבו מצאו תומכי הכאוס בביולוגיה כר לפעילותם. מה עשוי לגרום לאבולוציה, והאם אין סתירה בין דטרמיניזם לאבולוציה?
גם דטרמיניסט מושבע אינו יכול להתעלם 41מהאפשרות שמצב ״כאוטי״ בטבע עלול לגרום לסטייה מקרית של המערכת החיה, מהמסלול הדטרמיניסטי הראשוני שלה. מתי נתייחס לשינוי כזה כאל סטייה מיכולת החיזוי ומתי לא? ובניסוח אחר – מהו ה״קו האדום״ של הדטרמיניזם?
חשוב להבהיר, כי גם דטרמיניזם אינו מצב מוחלט, וכי גם בו קיימות דרגות שונות. מכיוון שהחיים לא נותרו בצורתם הראשונית, חייבים היו לקרות בהם שינויים גנטיים שהוליכו אותם, ובצורה מסודרת, מהצורות הראשוניות לצורות מפותחות יותר. לכאורה יש בכך סתירה: מחד גיסא יש לפנינו מערכת דטרמיניסטית; מאידך גיסא, זאת מערכת ה״מעכלת״ שינויים ומאפשרת אבולוציה.
אורגניזם חי מתוכנת לדכא שונות שאינה מתוכננת
לאירגון יציב וחזוי מראש יש יתרון. לכן, אורגינזמים חיים נוטים להישאר במידת האפשר בצורה היציבה והקבועה ביותר. ומה ביחס לשינויים האבולוציוניים הקטנים שהופיעו במהלך הדורות? אלה היו חייבים להיות מצומצמים מאוד, ובעוצמה הקטנה מעוצמת הסף, שיש בה כדי להפוך את תהליכי החיים לכאוס. קליטת שינויים במערכת החיה התאפשרה כל עוד לא עירערה את הבסיס הדטרמיניסטי של המשך קיומה, ובזכות ההגנה המובנית מפני הפרעות חיצוניות בלתי רצויות. בצורה בוטה אפשר לומר, שאורגניזם חי מתוכנת לדכא שונות שאינה מתוכננת. עם זאת, לא תמיד אנחנו יכולים לחזות את כיוונם של השינויים הקטנים הללו, וכיווני ההתפתחות האבולוציוניים היו ככל הנראה בלתי חזויים. זאת גם הסיבה, שיש ביולוגים הרואים בכאוס את התהליך המוביל באבולוציה. אבל, ברור שבגישה כזאת יש משום חוסר פרופורציה בהערכת ממדי השינויים שקרו בכל פעם: האם פיל, אשר פיתח רגליים קירחות כתוצאה משינוי גנטי (מקרי או אבולוציוני), חדל להיות פיל? מה צריכים להיות ממדי השינוי בצורתו של הפיל כדי שהוא יפסיק להיות פיל? יש אפוא להגדיר את גבולות השינויים שיכולים להיטמע במערכת חיה, שינויים אשר האורגניזם הקיים יכול לחזות אותם ואילו מעבר להם, נוצרת איכות חדשה, בעלת חוקיות שונה.
מבחינה זו, אבולוציה היא תהליך חיובי, המבטא צירופי מידע חדשים ויכולת להתמודד עם תנאים שונים. אבל אל נשכח, שצירופי המידע האלה יתקיימו רק אם הם יהיו מאורגנים, יתרמו להתאמת האורגניזם בסביבתו, וישמרו על יכולתם לחזור על עצמם. לפיכך, גם מנקודת השקפה זו אין החיים יכולים להתבסס על מקריות. אמנם אפשר להתייחס לאחדים מהשינויים האבולוציוניים כאל ״מחלה״, המתבטאת באירגון חדש, אבל בדיקה מדוקדקת מוכיחה שאפילו חלק ניכר מהשינויים הגנטיים הנראים בתאים חיים הם מתוכנתים. הם כוללים מוטציות מושרות ומכוונות, שיחלוף מכוון של קטעי כרומוזומים, טרנספוזונים2 מוגדרים, מוטציות המפוקחות על ידי גנים ספציפיים ומערכות תיקון דטרמיניסטיות.

גם אתרי ההצרפה (רקומבינציה)3 אינם מפוזרים באקראי על פני גדילי ה-DNA אלא מתרכזים ב״קבוצות מידע 42ספציפיות״. לפיכך, אפילו השינויים הגנטיים במערכות חיות, שנראו בעבר ככאוטיים, עשויים להיות דטרמיניסטיים.

ביטויים של פרקטליות ביצורים חיים: האם זה מקרי?
לעיתים קושרים בין ביטויים פרקטליים במערכות חיות לבין כאוס ומקריות. במבנים פרקטליים, צורות מסוימות חוזרות על עצמן בקני מידה שונים. מבחינה מתמטית הם מאופיינים על ידי ממד שבור, בעוד מבנים שאינם פרקטליים הם בעלי ממד שלם (ממד של קו הוא 1, ממד של משטח הוא 2 וממד של תיבה הוא 3- ראה יששכרי, מחשבות 66, אפריל 1994). כך, למשל, סידור העלעלים על גבי עלה, או אירגון מערכת כלי הדם בגוף האדם, מוזכרים בהקשר זה כדוגמה ל״מערכת פרקטלית-כאוטית״. אין ספק שניתן לתאר את מערכת כלי הדם בצורה גרפית בעלת ממדים פרקטליים. אבל גם אין ספק שלעולם היא לא היתה נוצרת, ולא היתה מתפקדת כמות שהיא, אילו המידע הגנטי הדרוש ליצירתה היה חסר כיוון וחסר ודאות. בהשאלה, ניתן אולי לתאר את בטנה התופחת של אשה הרה בצורה גרפית בעלת ממדים פרקטליים. אבל, מכאן ועד להצהרה שהריון הוא תהליך אקראי או כאוטי, המרחק הוא אינסופי. ייחודה של מערכת פרקטלית הוא ביכולתה לבטא את החוקיות שבתכונותיהם של מבנים, החוזרים על עצמם בקני מידה שונים. אלא שלכל תהליך כזה בטבע יש מגבלות הן כלפי מעלה והן כלפי מטה.
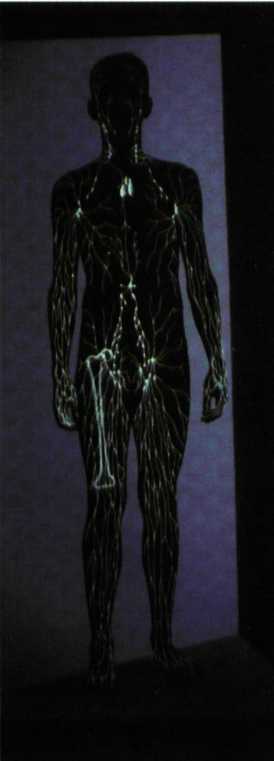
לכאורה, דמיון עצמי מוגבל מצד אחד במבנה האטום ומצד שני בממדי היקום. אבל, כאשר אנחנו עוסקים במערכות חיות, השינויים מוגבלים רק לתחום מצומצם של קני המידה, שבהם מרכיבי היסוד של המבנה או של המידע משמרים את אופיים, או שבהם האורגניזם יכול להמשיך לחיות. במשחקי מחשב ניתן להדגים מצבים שבהם ״דג״ בולע ״דג״, הבולע ״דג״, הבולע ״דג״ וכו׳. זוהי דוגמה מתמטית המראה שקנה המידה הולך וקטן למרות שהדג שומר על צורתו בטווח ממדים גדול. במערכת חיה לא ייתכן קיומם של דגים הקטנים מממד מסוים, משום שהם מאבדים אז את ה״דגיות״ שלהם, ולא ייתכן קיומם של דגים הגדולים מממד אחד, משום שהם ימותו מחוסר משאבים או מחוסר יכולת תנועה. אפשר לחזות שינויים בממדי הדגים אך רק בטווח מצומצם מאוד של קני מידה.

הזיהוי של פרקטליות וכאוס הוא מטעה מיסודו
דוגמה נוספת למגבלות של התיאור הפרקטלי היא בהסתעפות ענפים של עץ. הסתעפות כזאת מבטאת צורה פרקטלית, אבל רק במשך תקופה מצומצמת מאוד של מחזור חייו של העץ, ורק במספר קטן מאוד של ההסתעפויות של ענפיו. הדבר נעוץ במגבלות תכונותיו של האורגניזם הקרוי ״עץ״. לדוגמה, הסתעפות ענפים של עץ היא בדרך כלל אירוע שנתי. כלומר, בעץ בן מאה שנים, מספר ההסתעפויות האפשריות לאורכו של הגבעול הוא כמאה. אבל הדבר אינו מציאותי. התבוננות בעץ מוכיחה, שסעיפי הענפים שלו נעשים קצרים ודקים ככל שסדר ההסתעפויות עולה. אילו ההסתעפות העץ אכן היתה מבטאת מערכת פרקטלית, היא היתה חייבת ליצור סעיפי ענפים כה קטנים, עד כדי איבוד זהותם כאיברי צמח. כדי לתת דוגמה מספרית לכך נציין, שמשקלם של שני ענפי בת של עץ, או אורכם, קטן בממוצע פי 0.5 עד 0.77 ממשקלו של ענף האם (בהסתעפות דיכוטומית, היחס הוא 43כ-0.707, הערך הידוע בשם ״הערך של ליאונרדו דה וינצ׳י״, או ״ממד הזהב״). במלים אחרות, ירידת המשקל של הסעיפים היא כל כך משמעותית, עד שכבר לאחר 8-5 הסתעפויות נקבל במקום ענפים קבוצות תאים זעירות בלבד, אשד אין כל דמיון ביניהן ובין ענפי צמח. ענפי עץ עשויים אפוא ליצור דגם פרקטלי, אבל במגבלה של פחות מ-8 הסתעפויות. לאחר מכן משתנים לחלוטין אופיו של העץ וחוקיות ההסתעפות של ענפיו. העיקרון של ״נכון בכל קנה מידה״ הוא עיקרון חשוב ביותר בטבע, אבל מן הנאמר ברור, שלנתח את צורתם של אורגניזמים חיים בצורה פרקטלית ניתן רק בגבולות מצומצמים מאוד.
הנטייה לקשור בין פרקטליות וכאוס נבעה מן הרצון לחשוף תופעות של אי סדר במערכות שבהן המרכיבים הם בעלי ממדים שונים, אבל מחמת המורכבות של מערכות אלה תוארו רבות מהן כמערכות בלתי חזויות. ברור שיש בכך שגיאה. כאשר מערכת יוצרת מבנה פרקטלי, היא יוצרת מבנה מאורגן ובעל חוקיות. למשל, כאשר פתית שלג נוצר מסביב לגרעין גיבוש מסוים ויוצר מבנה גיאומטרי פרקטלי, פירושו של דבר שכל מולקולת מים שנוספה לגביש הראשוני, התחברה אליו דווקא במקום מסוים, בסדר מסוים והודות לכוחות מסוימים. מעצם מהותו, סדר כזה אשר חוזר על עצמו, אם כי בקני מידה שונים, אינו יכול להיות כאוטי. הדרישה ממצב כאוטי ש״תוצריו אינם ניתנים לחיזוי״ אינה ישימה במערכת פרקטלית, שהדמיון בין הרכיבים בה הוא מלא. לו יצירת שלג היתה תופעה כאוטית, לא היו נוצרים ״כוכבי השלג״ המוכרים, אלא היתה נוצרת מסה חסרת צורה מוגדרת וחסרת מבנה. מכאן שהזיהוי של פרקטליות וכאוס הוא מטעה מיסודו.

מוות: דטרמיניזם או כאוס?
קיומן של צורות מתקדמות של חיים התאפשר רק לאחר שהתפתחו בהם מנגנונים מוגדרים, המווסתים את התרבות התאים, את התמיינותם, ואת שינוי פעילותם עם הזמן. אין ספק שמנגנונים אלה הם מתוכנתים וחזויים מראש. אבל, לא רק החיים מתוכנתים בתאים חיים אלא גם המוות. באורגניזם חי חייב להיות בכל רגע ורגע מצב של שיווי משקל בין תאים נוצרים ובין תאים מתים. כדי להבטיח שבכל רקמה יישארו תאים רק במספר הדרוש לה, מתוכנתים חלק מהם ״להתאבד״.
מבחינת האורגניזם השלם, המוות ממלא תפקיד חיוני בהרחקת תאים בלתי רצויים. עם זאת, אם המוות יהיה מקרי ולא מתוכנן מראש (למשל על ידי גורם סביבתי, שאינו בשליטת האורגניזם), הוא עלול לפגוע בתאים חיוניים ולגרום בכך נזק לאורגניזם השלם. אחד המנגנונים החשובים בשמירת הקיום הביולוגי (בכל רמות האירגון) הוא מנגנון ההקרבה של הפרט לטובת הכלל – הקרבת תאים, הקרבת איברים או הקרבת אורגניזמים. לדוגמה, התאבדות של תאים של רקמת העור מקיימת את האורגניזם בכך שהיא קוטלת את התאים ה״כאוטיים״, תאים שהמערכת הגנטית שלהם איבדה את תיכנותה הראשוני כתוצאה מקרינת uv. מערכת דומה גורמת להרחקה מסודרת של תאים של מערכת העיכול שה-DNA44 שלהם נפגע מרעלנים. תופעת הנשירה בצמחים אינה אלא ״התאבדות מתוכננת״ של עלי העץ, לאחר שצברו במהלך התפתחותם חומרי עיכוב, מחלות ומזיקים ואיבדו מיעילותם.
איך יודעים תאים שהם מיותרים ושהגיע זמנם למות? ובכן, במערכות חיות התפתחה צורה מסודרת של מוות, הנקראת אפופטוזיס (apoptosis) או מוות מתוכנת (programmed cell death). זהו תהליך האחראי להרחקת תאים מסוימים. מוות מתוכנת מבוסס על מערכת גנטית שהתפתחה במהלך האבולוציה ושכנראה מצויה היום בכל בעלי החיים, החל בתולעים וכלה בבני אדם, ואף בצמחים שונים. בניגוד למוות ״רגיל״, השלב הראשון של אפופטוזיס אינו מתבטא בפגיעה בממברנות של התא המתאבד, אלא בהתמוטטות של הגרעין; בהפעלה של אנזימים מפרקי חומצות גרעין (אנדונוקליאזות), ובשבירת הכרומטין לקטעים קטנים, הנותנים ל-DNA צורה דמוית סולם. כל תא העשוי למות בצורה מתוכננת מגיב לאות מסוים, המפעיל את תהליך המוות. אבל הרי אות זה עלול להיקלט על ידי תאים שכנים רבים של האורגניזם. ובכן, גם בכך יש סדר, והתאים שאינם מתוכנתים למוות יגיבו על אותו אות תגובה שונה לגמרי. ניתן לחסום את ההתאבדות על ידי הפרעה בביטוי הגנים האחראים לכך. למשל טיפול ב-Actinomycin-D (מעכב תיעתוק) או ב-Cycloheximide (מעכב של תירגום) ״חונן״ תאים ה״נדונים למוות״ ומאפשר להם להמשיך ולתפקד כרגיל. הפרעת הביטוי של הגנים מונעת אפוא מהתאים הללו את הגורם הכימי הדרוש להתאבדותם. ובכן, גם המוות ה״נורמלי״ של תאים חיים אינו בהכרח מקרי.

לסיכום:
מערכות חיות בנויות ומתפקדות בסדר קבוע. הבנת החוקיות של סדר זה, תהיה אשר תהיה הסיבה ליצירתו, היא אחד האתגרים המעניינים ביותר במדעי החיים.
מערכות ביולוגיות הן ביסודן מערכת דטרמיניסטיות, המסתמכות על מערכת מידע משוכללת. לעיתים אין הדטרמיניזם נראה לנו בצורה מוחשית ובולטת, אבל גם במקרים של ספק אפשר לראות בכל שלב ושלב של חיי האורגניזם את התוצאות של ״עריכה״ מתוכננת וקפדנית להרחקת שגיאות מקריות. כאשר מערכת ביולוגית עוברת ממצב של ודאות למצב של אי ודאות, היא מפסיקה להתקיים כמערכת חיה.
מערכת פרקטלית היא מערכת דטדמיניסטית. ניתן להשתמש בניתוח פרקטלי כדי לתאר את מרכיביו של אורגניזם חי. אבל לניתוח כזה יש מגבלות תמורות מבחינת קני המידה ומבחינת החוקיות של המבנים שהיא באה לתאר.
יציבותו של הטבע היא תוצאה של כללי התנהגות נוקשים של כל מרכיביו. התבוננות בטבע כבמערכת של תוהו ובוהו היא שעשוע נחמד. אבל יש לזכור שהטבע, כפי שאנו מכירים אותו, כולל בתוכו גם מרכיבים חיים -דטרמיניסטיים. משום כך, קיומו של הטבע כיחידה אקולוגית מתפקדת איננו יכול להיות כאוטי.
ואסיים בציטוט של אותו הרעיון כפי שבוטא בספרו של מאיר שלו, ״כימים אחדים״:
״והכי חשוב, שיינפלד, גם באוכל כמו באהבה, זה הכללים. בבית שאין בו כללים, הגורל משתולל, המזל מתעכב והמקרה מבקר. אבל בבית שיש בו כללים, הגורל עושה את מה שאומרים לו, את המזל לא צריכים, והמקרה נשאר בחוץ, דופק וצועק ולא יכול להיכנס״. ■
ספרות רקע:
גליק, ג׳יימס (1991). כאוס. מדע חדש נוצר. ספרית מעריב. תל אביב.
דוקינס, ר. (ו99ו). הגן האנוכיי. הוצאת דביר. תל אביב. ויץ, י. (1994). הרצאה במסגרת אופקים חדשניים בביולוגיה. אוניברסיטת תל־אביב.
זיו, יוסי (1993). שיחה עם ישעיהו ליבוביץ. מחשבות, 65: 23-6.
ינאי, צבי (1994). בעקבות המחשבות. פואטיקה, טובי ספר מוציאים לאור. רמת גן
יששכרי, דוד (1994). על תכונותיו הפרקטליות של הזמן. מחשבות, 66: 17-6.
ליבוביץ, ישעיהו (1994). הרצאה במסגרת אופקים חדשניים בביולוגיה. אוניברסיטת תל אביב

