





הקדמה:
ב-1975 ראה אור ספר עב-כרס, שהדיו מתגלגלים עד היום: ״סוציוביולוגיה – הסינתיזה החדשה״. מחברו, אדוארד או’ וילסון, היה מוכר בחוגי המקצוע כחוקר רציני שהתמחה בעיקר בחקר ההתנהגויות החברתיות של חרקים. אלא שבספרו החרש הרחיק לכת הרבה מעבר לתאור שיטתי ומדוייק של מנהגים והתנהגויות של בעלי-חיים. המונח החדש שטבע, סוציוביולוגיה, בא להצביע על הבסיס הביולוגי שביסוד ההתנהגויות החברתיות של בעלי-החיים לרבות האדם.
למעשה, וילסון צופה כי בעתיד הלא רחוק יהפכו הסוציולוגיה, הפסיכולוגיה, האנתרופולוגיה, הביולוגיה של אוכלוסיות, הגנטיקה האתולוגיה ועוד, לענפים של הסוציוביולוגיה. מיזוג הסוציוביולוגיה עם הנוירופיסיולוגיה, מעריך וילסון, עשוי לתת הסבר מדוע אנו מעדיפים במצבים מסויימים הכרעות מוסריות מסויימות על פני אחרות.
כל זה בגדר הערכה בלבד. לפי שעה מסתפק וילסון בקביעה שחוט אחד מוביל מהתנהגותן של מושבות טרמיטים, המקיימות דרגה גבוהה של שתפנות חברתית, להתנהגות החברתית של האדם.
בצטטו את אלברט קאמי, כי ״חתירה לגבהים דיה למלא ליבו של אדם״, טוען וילסון שאמירה זאת חסרה משמעות, אלא אם היא נבחנת לאורה של תורת האבולוציה. התודעה משועבדת ומעוצבת ע״י מרכזי הבקרה הרגשיים בהיפותלמוס ובמערכת הלימבית של המוח, כותב וילסון. מרכזים אלה, המציפים את תודעתנו ברגשי שנאה, אהבה, אשמה, פחד ועוד, משמשים את הפילוסופים בבואם לנסח אמות מידה של טוב ורע, אך אין לשכוח שמרכזים אלה הם תוצרי האבולוציה, והעובדה שהבירור הטבעי לא סילקם, כי אם הפיצם בכל המין האנושי, פירושה שיחסי הגומלין בין רגשות שונים ואף נוגדים אלה חיוניים להשרדותם ולהתפשטותם של הגנים האחראים ליצירתם ולתפקודם של מרכזים אלה. שהרי אלמלא כן, היו בעלי מרכזים אלה מפנים מקומם ליצורים אנושיים בעלי מרכזים מטיפוס שונה, אולי נטול רגשות.
המסקנה, איפוא, שהם חיוניים. אבל חיוניים למה? לא לאבטחת אושרו של האדם, אלא להעברה מירבית של הגנים הנושאים את תכנית הבניה של מרכזים אלה מדור לדור. שכן האושר או התוקפנות אינם חשובים כשלעצמם. חשיבותם נמדדת בתרומתם ליכולת השרדותם של הגנים ולכושר שכפולם. הוא הדין לגבי איברים, מנגנונים ביולוגיים ותכונות התנהגותיות של האורגניזמים השונים: חשיבותם אינה בתרומתם לכושר ההשרדות של הפרט כגרסתו של דארווין, אלא ביכולתם לשמר את הגנים שלו ולהבטיח להם תפוצה מירבית.
לכאורה ההבדל בין השניים סמנטי, שהרי ככל שהפרט חי ומתקיים זמן רב יותר, הוא יכול להוליד מספר רב יותר של צאצאים. ולהיפך – כל ארוע3 המביא למותו או פוגם בכושר פריונו מצמצם הלכה למעשה את כמות הגנים שהוא יכול בכוח להפיץ.
אלא שעובדות הטבע אינן פועלות בהכרח עפ״י הגיון זה. דורי דורות של זכרים ממשפחת הברחשים נקטלים תיכף למעשה ההזדווגות ע״י בנות זוגם. מדוע לא הכחיד הבירור הטבעי מנהג תמוה זה, המבטל את סיכוייו של הזכר להקים לעצמו דורות נוספים של צאצאים נושאי הגנים שלו ? מדוע לא העדיף היווצרות מנגנונים המאפשרים לברחש להמלט מתגובתה הקטלנית של הנקבה? וכיצד נסביר את קיומם של מנגנונים גנטיים בקרב עכברים כיסנאים (חיות-כיס) מסויימים, אשר האקט המיני גורם להם שטפי דם פנימיים ומביא למותם? יתירה מזאת, אם השרדות הפרט היא-היא שעומדת ביסוד דחף הקיום, אזי הנטייה האגואיסטית – כלומר העמדת האינטרס הפרטי מעל ולפני הכל – צריכה לשרור בכל עולם החי. כך, למשל, נטייתם של שחפים שחורי הראש לטרוף את גוזלי שכניהם צריכה להיות דגם התנהגותי כולל, שהרי בכך הם משיגים ארוחה דשנת קלוריות ללא מאמץ. ומדוע לא לאמץ את מנהגן של נקבות גמל שלמה והצרצר הירושלמי, הטורפות את בני זוגן לאחר ההזדווגות? או אף להגדיל ולעשות כמעשה היבחוש הגדל בתוך גוף אמו וטורף אותה מבפנים? אבל לצד צורות התנהגות כעין אלה רבים בעלי החיים בטבע המסכנים – ואף מקריבים – חייהם כדי לסייע ולהציל את צאצאיהם ואת שאיריהם הקרובים והרחוקים כאחד.
כיצד נסביר תופעות אלה? מה מניע צפורים מסויימות המקננות על הקרקע, להתחזות כפצועות בהתקרב טורף למשכנן ולדדות לפניו בכנף שמוטה כדי למשוך אליהן את תשומת ליבו ולהרחיקו מהקן? מה דוחף פרטים מסויימים בלהקות צפורים להזהיר בקולי-קולות את חבריהם בהתקרב נץ ולמשוך בכך את תשומת ליבו אליהם? ומה גורם לדבורים חסרות עוקץ (החיות באזור הטרופי) להתנפל בשצף-קצף על אויבן בהתקפת התאבדות (ראשן נקרע מעליהן), או של דבורות הדבש להותיר את עוקצן, על קרביהן, בגוף אויבן? מה מביא טרמיטים לרסק עצמם בגוף אויביהם באמצעות בלוטות נפץ, או להניע את פועלות הדבורים בעתות מחסור לכלות את חלבון רקמותיהן כדי להזין בו את הרימות בכוורת?
תופעות אלה, המפגינות תכונות אלטרואיסטיות מובהקות, טרדו את מנוחתו של דארווין והביאו אותו להגדירן כבעיה ״פטאלית״ לתורתו. שכן, עפ״י תפישתו, רק תכונה מועילה, שתורמת להשרדות הפרט, יכולה להשתמר ולהתפשט באוכלוסיית המין כולו. מכך נובע שתכונה אלטרואיסטית צריכה להתמעט בכל דור ולהתחסל כליל. שכן נושאי תכונת ההקרבה העצמית ניספים מהר ומוקדם יותר מפרטים אגואיסטים, ולפיכך גם סיכוייהם להוריש תכונה זאת לצאצאיהם קטנים יותר.
דארווין ניסה להחלץ מפראדוקס זה באמצעות רעיון הבירור הטבעי, הפועל ברמות המשפחה והפרט גם יחד. דהיינו, התכונה האלטרואיסטית תורמת לקיומה של הקהילה בכללותה ומקנה לה יתרון על-פני קהילה המורכבת מפרטים אנוכיים; מאחר שכך נהנית תכונה מוזרה זאת מעידודו של הבירור הטבעי.
היה זה פתרון דחוק למדי, שפתח פתח לפרשנויות מעורפלות על הבירור הטבעי הפועל לטובת הגזע והמין. אין זה מקרה, לכן, כי הפתרון שאומץ ע״י הסוציוביולוגיה, המציע את הגנים כשדה פעולה של הבירור הטבעי, עלה ופרח סביב שאלת האלטרואיזם.
אבל לפני שנרחיב את הדיון בנושא זה, נחזור לשעה קלה ללידת הדארוויניזם, כדי לעמוד – ולו במקצת על משמעותו של אחד מציוני הדרך החשובים ביותר בהתפתחות המדע.

לידת הדארוויניזם
צ׳ארלס דארווין נולד ב-1809, בשנה בה פירסם חוקר הטבע הנודע, ז׳אן באטיסט למארק, את ספרו על הפילוסופיה הזאולוגית. ב-1831, משהשלים לימודיו באוניברסיטאות אדינבורג וקמברידג’, הוצע לדארווין לעלות כגאולוג וחוקר טבע על פריגטת המחקר ״ביגל״, שעמדה לצאת למסע מיפוי בן 5 שנים לאורך חופי פטגוניה, טיירה דל-פואגי, צ׳ילי, פרו ומספר איים באוקיינוס השקט.
קטע המסע שהשפיע ביותר על גיבוש תורתו היה הביקור באיי גלאפגוס, השוכנים באזור קו המשווה, כ-1000 ק״מ ממערב לחופי אקוואדור. דארווין מנה באיי גלאפגוס 26 סוגים של צפורי יבשה (פרושים), שמתוכם 23 נראו כמינים נפרדים. מה שהפתיע אותו ביותר היתה העובדה שלמרות היותם מינים מקומיים דמו דמיון מופלג למיני צפורים החיות בדרום אמריקה, מרחק של 1000 ק״מ משם. תופעה זאת כה הרשימה אותו עד שהביאה אותו לציין ביומנו, כי היא קרבה אותו ל״סוד הסודות – להופעה הראשונה של יצורים חדשים על-פני כדור הארץ״. תופעה זאת וגילויים אחרים שנקרו לפניו בשמורות הטבע הקסומות של איי גלאפגוס, הביאו את דארווין לחשוב במונחים של אבולוציה באמצעות שינויים מיזערים ומצטברים.
דארווין שב ממסעו עם חלקים מרכזיים של תורתו, אך לכדי גיבושם לתפישה כוללת הגיע רק ב-1838, עם שנקלע לידיו ספרו של תומס רוברט מלטוס (1834-1766) על היחס בין גודלה של אוכלוסיה למשאבי המזון. הנחתו המרכזית של מלטוס היתה כי בעוד שייצור המזון גדל בטור אריתמטי (12,10,8,6,4,2, וכו׳) נוטה האוכלוסיה לגדול בטור גאומטרי (64,32,16,8,4,2 וכו׳). אי-לכך, מה שמבטיח לאוכלוסיה אמצעי קיום הוא שיעור תמותה גבוה מחמת רעב, מחלות ומלחמות, המקזזים את צמיחת היתר של האוכלוסיה ביחס למזון ושומרים על האיזון הנחוץ בין השניים.
התיזה של מלטוס נפלה על קרקע פוריה, שכן דארווין התהלך שנים רבות תחת הרושם הכביר של תהליך ההשמדה ההדדית בטבע ושל שיאי הפריון והתמותה הפוקדים את המינים השונים במחזוריות סדירה. ומתוך חוקיות זאת קמה והתייצבה לפניו אחת החוליות החשובות של תורתו, כפי שהוא מעיד עליה ביומנו: ״ומהיותי מוכן היטב להעריך, מתוך תצפיות ממושכות על הרגליהם של חיות וצמחים, את מאבק הקיום המתנהל בכל מקום, התברר לי לפתע שבתנאים כאלה שינויים חיוביים יהיו נוטים להשתמר ואילו השליליים להכחד. התוצאה תהיה היווצרות מינים חדשים״.

ספרו של מלטוס סיפק איפוא לדארווין את המכאניזם של הבירור הטבעי: השינוי החיובי ישתמר בפרט, אשר יפיצו באוכלוסיה באמצעות צאצאיו; ואילו הפרט הנושא שינוי שלילי יפסיד במאבק עם בני מינו על המזון והנקבות, עקב זאת יהיה פגיע יותר למפגעים של הסביבה העויינת ומספר הצאצאים שינומיד יהיה קטן יותר, כך שבמהלך מספר דורות תיכחד שושלתו כליל.
מכאניזם קר ונטול רחמים זה של הטבע השפיע עמוקות על דארווין. ״אין דבר קשה יותר״, כותב דארווין בזכרונותיו, ״מאשר לזכור ללא הפוגה את דבר קיומו של מאבק הקיום האוניברסלי״. ובהמשך הוא מוסיף: ״לנוכח זמרת הצפורים אנו נוטים לשכוח שרובם חיים על חרקים ולפיכך משמידים ללא הרף חיים; אנו שוכחים שצפורי שיר אלה מושמדים ע״י טורפים״.
מוצא המינים
אף שהחזיק בידיו את כל מרכיבי תורתו, היסס דארווין להוציאה לאור. יותר מעשרים שנה נדרשו לו למן מסעו ב״ביגל״ כדי לפרסם ברבים את תפישתו על השתלשלות האורגניזמים השונים ממוצא משותף, ועל מקומו של הבירור הטבעי הפועל באמצעות4 השרדות הכשירים, כגורם מרכזי ביצירת מינים חדשים.
ידידיו דחקו בו לפרסם את תורתו, מה גם שהוגים וחוקרים אחרים, כמו רוברט צ’מברס, נתנו מהלכים להשקפות דומות על השתלשלות בעלי החיים במהלך האבולוציה. אין זאת שהדחיפה האחרונה להוצאת ספרו לאור באה מכתב-יד של אלפרד ואלס ראסל שהגיע לידיו, אשר הצביע, בין השאר, על השינויים החלים בבעלי החיים כבסיס ליצירת מינים חדשים. ואמנם, ב-1858 שיגרו ואלס ודארווין מאמר משותף על תורת האבולוציה לאגודה הלינארית, אלא שמאמרם לא עורר תשומת לב מיוחדת. ואז, ב-1859, הוציא סוף-סוף דארווין לאור את ספרו על מוצא המינים ובכך ניתן האות למהומה הגדולה שהסעירה את הקהילה המדעית ואת הציבור הרחב גם יחד.

המסקנה הנגזרת מספרו (אשר זכתה לאמירה מפורשת בספרו המאוחר משנת 1871, ״מוצא האדם״), כי אבות האדם היו קופים, עוררה נחשול אדיר של תגובות נזעמות. לפופולריות מיוחדת זכתה הערכתו של הבישוף סמואל וילברפורס, אשר בכנס פומבי ב-1860 שאל האם כל מי שמסכים להכיר בקוף כבסבו חייב להכיר גם בקופה כסבתו. תגובות נזעמות אלה מלמדות כי בני דורו של דארווין היו קרובים יותר לתפישתו של קרל פון לינאוס מאשר לזו של דארווין. לינאוס, האיש שהגה את סיווגם ההירארכי של בעלי החיים למינים, לסוגים, למשפחות, לסדרות, למחלקות ולמערכות – גרס כי מספר המינים הקיימים קבוע ועומד מיום שאלוהים בראם, ומכאן: לא יתכן שקיימים בימינו יותר מינים של בעלי חיים מאשר ברגע הבריאה.
עם זאת, מושג האבולוציה לא היה חדש ובוודאי לא זר לאוזן. כבר ב-1559 כתב הנזיר יוהנס בוטהו מאמר, המנתח את צורתה ומידותיה של תיבת נוח ומקיש מכך שלא היתה יכולה להכיל את כל אוכלוסיית המינים השוכנת על-פני כדור-הארץ. מכך גזר שמינים רבים וחדשים, החיים בעת המודרנית, התפתחו מנוסעיה המקוריים של התיבה.
ב-1687 כתב הפילוסוף לייבניץ, כי יצורים חיים משתלשלים זה מזה באמצעות שינויים מזעריים. בדרך זאת נוצר סולם רציף של חיים, שבו השלבים המייצגים את הפרטים נבדלים זה מזה מבחינה כמותית ולא איכותית.

למעשה, המונח אבולוציה הוצע לראשונה ב-1744, ע״י הביולוג הגרמני אלברט פון הלר, במטרה לתאר את התפתחות העובר מיצור קדם-עוברי: ״הומונקולי״. פון הלר גרס, שההומונקולי הזעיר ארוז בצורתו המוגמרת כבר בביצית או בתא הזרע. על-פי תפישתו, כל בני אנוש החיים עמנו התקיימו כבר כהומונקולים בתאי המין של אדם וחווה.
הוגה דעות נכבד שהצביע על אפשרות של מוצא משותף מתוך הדמיון המורפולוגי הניכר בין צורות החיים השונות, היה קאנט, אף שהגדיר רעיון זה כ״הרפתקאה נועזת של המחשבה״.
בתחום האמפירי אפשר להצביע על שני גילויים סנסציוניים, אשר עשו נפשות לרעיון האבולוציה. ב-1739 גילה טרמבליי את ההידרה, רב-תאי זעיר החי במים מתוקים, המסוגל להתרבות ע״י הוצאת חוטרים, משל היה צמח, ולפיכך הוצג בציבור כחוליה המקשרת בין עולם הצומח לעולם החי. אבל דומה שהעדות המרעישה והמשכנעת ביותר בכיוון זה היה גילוי המאובן של הארכאופטריקס (Archaeopteryx) יצור קדום בעל זנב-עצם ושניים כשל זוחל, אך בעל נוצות ומקור כשל עוף. גילוי המאובן המרתק הזה בשנת 1861 היה לרבים הוכחה ניצחת להתפתחות העופות מהזוחלים.
למארק והנחלת התכונות הנרכשות
אכן, דומה כי המאה ה-19 היתה בשלה לרעיון האבולוציה. כבר בראשיתה של אותה מאה התייצב מחנה האבולוציוניסטים, בראשותם של ארסמוס דארווין (1802-1731) – סבו של צ׳ארלס דארווין, ושל ז׳אן באטיסט למארק (1829-1744), שתורתו חוזרת ומגיחה כרוח רפאים בתוך דיוני האבולוציה של ימינו-אנו.5 ואמנם, בצל שני אישים סמכותיים אלה, למארק ודארווין הסב, הגיע המאבק בין חסידי הבריאה לחסידי ההתפתחות לשיאו, עם פרסום ספרו של התאולוג ויליאם פאליי, שניסה להזים, אחת ולתמיד – באמצעות המשל של השעון – את הרעיונות ״הנואלים״ של האבולוציה. כל המתבונן בפעולות המתואמות של הקפיצים והגלגלים המניעים את מחוגי השעון, טען פאליי, לא יכול שלא לעמוד על התכנון שמאחוריו, קל וחומר העין האנושית. ממורכבותה ומתפקודה של העין אנו למדים על קיומו של מתכנן, המודע היטב לחוקים הנעלמים ביותר של האופטיקה. במשלו התכוון פאליי, כמובן, להצביע על כך שאיברים מורכבים – לא כל שכן אורגניזמים שלמים – אינם יכולים להיווצר בצורה אקראית מעצמם; שכן הם מגשימים מטרה שקבע להם המתכנן – במקרה זה בורא העולם.

נראה שלפאליי היתה סיבה טובה לרתום את כשרונו הרטורי לעזרת הבריאה האלהית. שכן הרעיונות ההתפתחותיים, שערם נלחם, התגבשו כעבור מספר שנים לתאוריה שלמה ועקיבה בספר שפירסם למארק ב-1809. למארק תאר את המכאניזמים הטבעיים העומדים ביסוד התפתחות החיים, מאורגניזמים נחותים למפותחים, ללא התערבותה והכוונתה של ההשגחה העליונה.
ביסוד התורה ההתפתחותית של למארק עמדו 4 עקרונות: הדחף הפנימי לשלמות, יכולתו של אורגניזם להסתגל לסביבתו, התקיימות יצירה ספונטנית של חיים וכושר ההורשה של תכונות נרכשות מהורים לצאצאים.
למארק גרס שאיבר כלשהו של בעל-חיים יכול להשתנות ולהתפתח במרוצת הדורות בגין שימוש יתר בו, או להתנוון ולהעלם אם אינו מועיל ולא נעשה בו שימוש. למארק ראה בדחף ובכורח הקיומיים את המנגנון העיקרי של השינוי. למשל, הג׳ירפה פיתחה צוואר ארוך עקב התדלדלות המזון על הקרקע, ומכאן הכורח הקיומי שניצב לפניה להגיע לעלים שבצמרות העצים. במילים אחרות, למארק – בניגוד קוטבי לתפישה שעתידה להבשיל אצל דארווין – האמין בקיומה של הסתגלות ישירה ואקטיבית של האורגניזם כלפי הסביבה. דהיינו, ההכרח להסתגל לסביבה לצרכי הקיום יוצר לחץ על פיתוח צוואר ארוך יותר (לג׳יראפה), שרירי רגליים חזקים יותר (לאיילה) או ניבים חדים יותר (לנמר).
מבחינה זאת, טענת הלעג עד תורת למארק, כי לאבות יהודים נולדים תינוקות בעלי ערלה, למרות שלהוריהם ולהורי-הוריהם עשו ברית-מילה, אינה ממין הענין. שכן כריתת הערלה לא מייצגת צורך קיומי ואיננה תוצאה של לחץ סביבתי, אלא של מנהג פולחני.

תורת למארק קנתה לה בראשיתה חסידים רבים, אך גם לאחר שנדחתה רשמית שבה ועלתה באצטלאות שונות בגרסאות המגוונות של החשיבה האבולוציונית. דארווין עצמו, אשר תורתו מוצגת כאנטי-תיזה לתפישה הלמארקיסטית, צידד בפרסומיו המאוחרים באפשרותה של הנחלה תורשתית של תכונות התנהגותיות ופיסיולוגיות נרכשות בקרב צמחים ובעלי חיים מבוייתים.
כיוון אחד למידע
למעשה, היתה זאת עבודתו של אוגוסט ויסמן (1914-1834), אשר הנחיתה מכה ניצחת לרעיון של הורשת תכונות נרכשות. ויסמן קבע כי תאי המין הזכריים והנקביים, האחראיים להולדת צאצאים, כמעט ואינם מושפעים מלחצים סביבתיים (שינויי טמפרטורה, מחסור במזון וכוי) או משינויים מתחוללים בגוף (פציעה, נכות וכו׳). ויסמן הבחין בין שני תהליכים המאפיינים את התחלקות הביצית ואת התמיינות התאים לאיברים השונים של הגוף (התמיינות לתאי כבד, עצב, עור וכו׳): תהליך אחד יוצר את תאי הגוף (soma) המושפעים מהסביבה ומגיבים כלפיה, אך סופם להעלם עם מותו של בעל הגוף. התהליך השני יוצר את תאי המין (המכילים רק את מחצית הכרומוסומים), שאינם מושפעים משינויים בסביבה. תוכנם של תאי המין עובר בתורשה מרור לדור ולפיכך מסוגלים להיות נצחיים.
כיום מזוהה השוני בין שני התהליכים במונח שטבע פרנסיס קריק (מי שפיענח עם ג׳ימס ואטסון את מבנה הצופן הגנטי) – ״הדוגמה המרכזית״ של הגנטיקה המולקולרית. דוגמה זאת קובעת, שהמידע התורשתי לעולם נע בכיוון אחד: ממולקולות הדנ״א שבגרעין לריבוסומים שבתא, המייצרים עפ״י מידע זה את החלבונים והאנזימים הדרושים לתיפקודו ולבנייתו של התא. לעולם אין המידע זורם מהתא ומחלבוניו אל החומר הגנטי שבגרעין. מכאן, שינוי המתחולל בתא אינו יכול להשפיע על תוכן החומר הגנטי; לעומת זאת, כל שינוי בדנ״א משנה את המידע הכלול בו ולפיכך, משיגיע לריבוסומים, ייצר חלבונים ואנזימים שונים. דהיינו – עקב השינוי בדנ״א יתפקד התא בצורה אחרת.
כיוון חד-סטרי זה של זרימת המידע בתא מבטל, בעקרון, את אפשרותה של תורת למארק. תינוק של נפח לא יוולד עם זרועות שריריות – גם אם אביו הוא דור עשירי של נפחים, שכן שרירים הם תוצאה של שינויים שחוללה הסביבה בתאי הגוף ולא בחומר הגנטי שלהם. תינוק אסקימוסי לא נולד עם פרווה, למרות הלחץ הסביבתי בן עשרות דורות. אם יוולד תינוק שעיר יהיה זה בגלל שינוי גנטי, והתפשטותו של שינוי זה באוכלוסיית האסקימוסים תתנהל עפ״י העקרונות הדארוויניסטים ולא הלמארקיסטים.
אך, כאמור, הרעיון הלמארקיסטי מסרב עדיין למות. מחקרים בתחום המיקרוביולוגיה המודרנית מצביעים על כך שהדוגמה המרכזית אינה מוחלטת ונחרצת.
כך, למשל, יש ראיות כי במקרים מסויימים הרנ״א בתא לא רק מעביר מידע מהדנ״א לאברונים המייצרים חלבונים, כי אם גם מזין חזרה מידע לדנ״א. מחקרים אחרים מראים על נוכחותם של גורמים תורשתיים בציטופלסמה של התא (כלומר, מחוץ לכרומוסומים), הנתונים לשינויים בהשפעת גורמים חיצוניים. לאחרונה הציג אימונולוג אוסטרלי, טד סטיל, תאוריה מעניינת בדבר היווצרות נוגדנים בגוף, העושה שימוש מעורב בעקרונות דארוויניסטים ולמארקיסטים.
לפי סטיל, תאי הלימפוציטים מפתחים נוגדנים ספציפיים עד האנטיגנים החודרים לגוף, ולאחר שפיתחו נוגדנים אלה עובר מידע זה אל הדנ״א למשמרת קבע ומונחל בתורשה לצאצאים.
המנגנונים האקראיים והדטרמיניסטים של האבולוציה
אבל נחזור למהומה הגדולה שהתחוללה בעקבות ספרו של דארווין. כאמור, הציבור התקומם נגד התפישה שהופעת האדם כפופה לאותם תהליכים החלים על כלל בעלי החיים.
במישור פחות פופולרי עוררה תורתו של דארווין מחאה פילוסופית בגלל המנגנון האקראי ומשולל התכלית של השינויים. לא ייתכן, טענו המתעדים, שהארמוניה המופלאה של הטבע והמבנה התואם והמשוכלל של האורגניזמים השונים, הם תוצאה של מקריות ואקראיות. זאת ועוד, באבולוציה הדארוויניסטית איו מקום או תפקיד לנפש. תורה זאת היא כל-כולה פועל יוצא של חוקים ועקרונות מטריאליסטים מובהקים. שלא כבתפישה הלמארקיסטית, לפיה מגיב האורגניזם לסביבה באורח דינמי, על-פי דארווין מתחוללים השינויים באורח עיוור ופאסיבי, ללא תלות ב״רצונו״ של האורגניזם, ודינם נחרץ לשבט או לחסד ע״י המנגנון הדטרמיניסטי של הבירור הטבעי – על-פי התאמתם לסביבה. בין שהותיר דארווין מקום לאלהים כיוצר חוקי האבולוציה והביולוגיה, ובין שדחק רגליו לחלוטין, תורתו היא ביסודה מטריאליסטית – נסיבתית וניתנת לרדוקציה למנגנונים ביולוגיים גרידא. מבחינה זאת היתה תורתו פרובוקטיבית כמו תגובתו המפורסמת של לאפלס לשאלה שהציג לו נפוליון היכן מקם במערכת הפיסיקלית שלו את בורא העולם. לאפלס השיב שלא היה לו צורך בהיפותיזה זאת.6 בתימצות רב ניתן לאפיין את תורתו של דארווין למספר עקרונות מרכזיים:
למינים השונים בקרב המשפחות השונות מוצא משותף. למשל – מוצא החרקים מאב משותף וכך גם העופות, הזוחלים והיונקים. ומכאן, ניתן להמשיך קו מחשבה זה ולגזור מוצא אחד משותף לכל האורגניזמים החיים בעולמנו. סיכוייו של פרט להשרד ולהתרבות תלויים במידת הסתגלותו לסביבה שהוא חי בה.
כל האורגניזמים בטבע הם בעלי כושר התרבות אדיר. רק אחוז מיזערי של כושר זה מתבטא בפועל, עקב התמותה הגבוהה, הנובעת ממשאבי המזון המוגבלים של הסביבה, מתנאים עויינים ומטורפים.
הפרטים השורדים הם בדרך כלל אלה המיטיבים להתחמק מטורפים והמצליחים יותר להשיג מזון ומחסה.
מינים חדשים נוצרים כאשר מציאות גאולוגית (כמו נדידת היבשות) או גאוגרפית (כמו שרשרת הרים גבוהה או ימה רחבה) מפלגת אוכלוסיה מסויימת ומונעת מגע בין שתי האוכלוסיות הבנות. במרוצת הזמן, בעקבות מוטציות, לחצי הסתגלות לסביבה החדשה והבירור הטבעי, צוברות האוכלוסיות הבנות שוני גנטי גדל והולך, עד לנקודה שבה הזדווגות בין שני פרטים מאוכלוסיות שונות תהיה עקרה או תוליד צאצאים עקרים.
האבולוציה מתנהלת בכוח שלושה מנגנונים עיקריים: מנגנון התורשה (החומר הגנטי), מנגנון המוטציות (שינויים החלים בחומר הגנטי) – המאפשר את האבולוציה, ומנגנון הבירור הטבעי – המבטיח את כיווניה של האבולוציה.
המנגנון הראשון מבטיח את המשכיותן והשתמרותן של הצורות והתכונות של האורגניזמים. המנגנון השני קובע כי האורגניזמים בטבע עוברים בהתמדה שינויים מזעריים. שינויים אלה מצטברים ונשמרים באורגניזמים ע״י המנגנון הראשון, האחראי גם להנחלתם בתורשה מדור לדור. המנגנון השלישי חורץ דינן של מוטציות אלו – מי להשרדות ומי להכחדה, עפ״י מידת מועילותן למלחמת הקיום ולמאבק המיני של האורגניזם וע״י תרומתן לכושר הסתגלותו ומידת התאמתו לתנאים המשתנים של סביבתו. מכאן, האבולוציה היא תוצאה של הצטברות הדרגתית של שינויים יתרוניים אלה, אשר באמצעות הרבייה והבירור הטבעי מתפשטים בהדרגה בין הפרטים והופכים את בעליהם לרוב המכריע והמצליח באוכלוסיה המורכבת מבני מינם.
מכאן גם ברור מדוע מנגנונים אלה קשורים זה בזה: מנגנון מוטציות ללא המסננת של הבירור הטבעי, היה מונע התפתחותם של מכאניזמים (ותכונות) ירודים ונחשלים למתקדמים ומשוכללים יותר, ולהיפך: מנגנון של בירור טבעי ללא מוטציות היה מקפיא את החיים כבר בשלב ההתחלי של המולקולות האורגניות הראשונות או מכחיד אותם בעקבות שינוי קיצוני של תנאי הסביבה.
האפונים של גרגור מנדל
המעניין הוא שדארווין ניסח את עקרון הפעולה של שני מנגנונים אלה בלי לדעת דבר על הדרך שבה מתרחשים ומצטברים השינויים המיזעריים ועל האופן שהם עוברים בתורשה מדור לדור. במרוצת השנים אימץ נוסח מעורפל בדבר התלכדות תכונות ההורים בתוך צאצאיהם, אלא שנוסח זה לא עלה בקנה אחד עם תופעות מובהקות של הטבע, שבהן תכונות פיסיולוגיות מסויימות עוברות בשלמותן מאחד ההורים לצאצאיו בלי להתלכד עם התכונות של ההורה השני. דארווין התחבט בסוגיה זאת עד סוף ימיו כמעט, אך ללא הועיל. אין זאת שהגורל החליט להתל בדארווין, שכן, בעוד הוא תר אחר מנגנון נסתר וחמקמק זה של האבולוציה, פורסם (ב-1865) בכתב-עת אוסטרי מקומי וכמעט אלמוני, מאמר בחתימתו של אח אוגוסטיני, גרגור מנדל, שנשא את השם ״ניסויים בזני כלאים של צמחים״, ובו דיווח על ממצאים שהעלה בהכלאת אפונה ירוקה. ממצאיו הראו שתכונות מסויימות עוברות מדור לדור בשלמותן ובכך חשף, למעשה, את החוקים הבסיסיים של התורשה הגנטית.
מנדל התרכז בניסוייו בשבע תכונות שונות של האפון ותרמילו: גודל, צורה, צבע, חלקות הקליפה ועוד. ע״י הכלאת אפון קמוט עם אפון חלק, או אפון צהוב עם אפון ירוק, מצא כי תכונות אלו מופיעות בשלמותן בקרב הצאצאים. כלומר, לעולם לא קיבל אפון חצי צהוב וחצי ירוק, או אפון מקומט למחצה, אלא אפון כולו צהוב או כולו ירוק וכיו״ב. משמע, התכונות שבדק מנדל לא התלכדו ולא התמזגו אצל הצאצאים כפי שגרס דארווין, כי אם עברו אליהם בשלמותן. עוד נוכח מנדל כי צאצאיהם הצהובים של הורים מעורבים, אם יוכלאו בינם לבין עצמם, לא יקימו דור של אפונים צהובים בלבד, כי אם דור מעורב המתפלג ל-75% אפונים צהובים ו-25% אפונים ירוקים. הסיבה להתפלגות ביחס זה של 1:3 הובנה סופית בשנות ה-20 של המאה שלנו, עם פרסום תורת הגנטיקה. מנדל עצמו לא הבין את המנגנון העומד ביסוד החוקים שגילה. הוא ייחס את המעבר השלם של התכונה מדור ההורים לצאצאים, לגורם תורשתי כלשהו, אך לא ידע מהו.
מעניין להעריך איזה השפעה היתה יכולה להיות לממצאיו על דארווין אילו הגיע מאמרו של מנדל לידיו. אמנם, העובדה שמנדל לא עמד על הזיקה של ממצאיו לתורתו של דארווין – שאת ספרו קרא אומרת דרשני. ובכל זאת, אין להוציא מכלל אפשרות שדארווין היה מגלה בממצאיו של מנדל את הסוכן הסמוי של השינויים המזעריים. כך או כך, אותו גורל שהיתל כדארווין עשה חסד עם מנדל, שכן לא במעט מזל ומקריות בחר בזני הורים הנבדלים זה מזה בתכונות המיוצגות בשלמותן ע״י גן אחד. הודות למקרה מוצלח זה חשף מנדל את חוקי התורשה הבסיסיים, וטבע, לראשונה, את המונחים דומיננטיות ורצסיביות, המיוחסים כיום לפעולת הגנים.
גילוי המנגנון הגנטי
ממצאיו של גרגור מנדל זכו להכרה מלאה ב-1900, עם גילוי מאמרו מחדש. בשנות ה-30 וה-40 של המאה שלנו, אוחדו המנדליזם והדארוויניזם לתורה הניאו-דארוויניסטית (או בכינויה האחר – התאוריה הסינתטית), שהתבססה על תפקיד הכרומוסומים (נושאי הגנים) בתהליך הרבייה המינית.
ההתפתחויות והתגליות בשטח הביולוגיה המולקולרית בעשרים השנים האחרונות, הוסיפו מימד חדש להבנת התהליכים האבולוציוניים. החדירה למבנה המולקולרי של החומר התורשתי, בעקבות פיענוח הצופן הגנטי של הדנ״א, גילתה כי שינוי בודד באחד מארבעת הנוקליאוטידים המרכיבים את מולקולות הדנ״א יכול להשפיע על התכונות המורשות לצאצאים.
למשל, מחלת אנמיה קשה, הפוקדת את תושבי ארצות אפריקה המשוונית, מאופיינת בכך שכדוריות הדם אצל נושאי המחלה מקבלות צורת חרמש (מכאן שם המחלה – אנמיה חרמשית), ומאבדות מכושר נשיאת החמצן. עקב גבשושיות הנוצרות על קרום התא – הן נתקעות בכלי הדם, עוצרות את זרימתו וגורמות לנמק של הרקמות.
מתברר שליקוי קטלני זה נובע משינוי של חומצה אמינית אחת מתוך שרשרת של 574 חומצות אמיניות המרכיבות את המולקולה של ההמוגלובין (החומר הקושר את החמצן לכדוריות הדם). והנה, נמצא שכדי לחולל שינוי זה, דהיינו כדי להחליף את החומצה האמינית התקינה באחרת, די לשנות נוקליאוטיד אחד מתוך רצף ארבעת הנוקליאוטידים המהווים את הא-ב של הדנ״א. ומכאן, יש ומוטציה אחת המתרחשת בהרכב הגנטי של הפרט (הגנוטיפ) מסוגלת לחולל שינוי משמעותי בתכונה עלית של הפרט (הפנוטיפ).
פיענוח הצופן הגנטי פתר את סודו של גורם התורשה המסתורי, וגם איפשר להרחיב ולהעמיק את המושג הרב-גוניות הגנטית (דהיינו, מגוון הגנים באוכלוסיה של מין נתון). רב-גוניות זאת, המשמשת לאבולוציה בסיס פעולה, יכולה לנבוע לא רק ממוטציות בגנים, שמקורן בגורם חיצוני-סביבתי ובטעות בתהליך השכפול של הדנ״א, היא גם תוצאה של תהליך השיחלוף וההרכבה מחדש של הכרומוסומים (של הזכר והנקבה) בתהליך יצירת תאי המין.
כדי להבין כיצד משתחלפים הגנים ואיזה תפקיד הם ממלאים בתהליך האבולוציה, הבה ננגיף מבט על תהליך הרבייה המינית.
על גנים ועל אללים
מולקולות הדנ״א ארוזות בגרעין התא בחוטים מעובים, הקרויים כרומוסומים, שכל אחד מהם נושא אלפי פיסות של דנ״א: גנים. הכרומוסומים מסודרים זוגות-זוגות ומספרם שונה ממין למין (לעכבר 40 כרומוסומים, לאדם 46, לתפוח-אדמה 48). 46 הכרומוסומים של האדם, למשל, מאורגנים ב-23 זוגות ומורכבים שווה בשווה מהגנים של האם ושל האב.
כללית אפשר לומר שהקוד הגנטי של כל תכונה גופנית שלנו רשום פעמיים – פעם בגן שירשנו מהאב ופעם בבן זוגו שירשנו מהאם. יש ושני הגנים המקבילים נושאים מידע זהה לחלוטין, ויש שהמידע זהה מבחינה פונקציונלית, אך ביטויו שונה. למשל, גן אחד נושא מידע ליצירת צבע עיניים כחול, ואילו מקבילו בכרומוסום הנגדי נושא מידע ליצירת צבע עיניים חום.
אם שני הגנים זהים בכל (במקרה שלנו – שניהם מכילים צופן לצבע כחול), נאמר על בעל החיים הנדון כי באתר מסויים זה שעל-פני הכרומוסום הוא הומוזיגוטי, ומכאן שצבע עיניו הוא בהכרח כחול. לעומת זאת, כאשר צמדי הגנים שבאתרים המקבילים נושאים מידע בביטוי שונה (למשל, גן אחד נושא מידע לצבע עינים כחול ובן זוגו מידע לצבע חום), הם קרויים אללים, ועל בעל החיים נאמר כי באתר מסויים זה הוא הטרוזיגוטי. צבע עיניו של פרט הטרוזיגוטי זה יקבע עפ״י הדומיננטיות של הגן: אם האלל הכחול הוא דומיננטי – יהיה צבע עיניו כחול, ואם להיפך – יהיה צבע עיניו חום.
אם כן, כיצד משרת מבנה זה של אללים את האבולוציה ואת הבירור הטבעי?
כידוע, כל התאים בגופנו – למעט תאי המין – מכילים בגרעיניהם מערכת זוגית של כרומוסומים (דיפלואידים), משמע: נושאים 46 כרומוסומים המאורגנים ב-23 זוגות. תאי המין, לעומת זאת,7 מכילים רק מערכת אחת (הפלואידים) בת 23 כרומוסומים.
הפחתה זאת מ-46 ל-23 כרומוסומים, מתרחשת בעקבות תהליך המיוזיס שעוברים תאי הרבייה: תא הרבייה הראשוני, שהוא דיפלואידי, משכפל את הכרומוסומים שלו (ל-92) ואחר-כך עובר פעמיים חלוקת הפחתה. בסיומו של תהליך זה מתקבלים 4 תאי מין הפלואידים, המכילים כל אחד 23 כרומוסומים בלבד. משלב זה ואילך עוברים תאי הרבייה הזכריים והנקביים מסלול שונה. אצל הזכריים – כל ארבעת הגרעינים ההפלואידים הופכים תאי זרע; ואילו אצל הנקביים – שלושה גרעינים הפלואידים מתנוונים ורק הרביעי הופך ביצית. כל אחד מ-23 הכרומוסומים המצויים בביצית או בתא הזרע עשויים פסיפס של קטעים מגנים של שני הוריו גם יחד, שהתערבבו ביניהם תוך כדי תהליך המיוזיס. ומאחר ששיחלוף זה של גנים בתהליך המיוזיס חוזר על עצמו בכל דור, יש לכל גן סיכוי להופיע באחד הצרופים ולעמוד במבחן הבירור הטבעי: אם הוא מועיל למאבק הקיום של הפרט ומשפר את התאמתו לסביבה – סיכוייו טובים לעבור לדור הצאצאים הבא; אם הוא מזיק ויש בו כדי לגרוע מכושר הסתגלותו של הפרט – גדולים סיכוייו להעלם תוך מספר דורות. התוצאה – ככל שינקפו הדורות יישרדו רק הגנים היתרוניים או הנייטרלים.
נחזור עתה לחוקי התורשה של מנדל ונעקוב כיצד הם משמשים את תהליך הבירור הטבעי.
המרור הטבעי והטלאים הצמריים
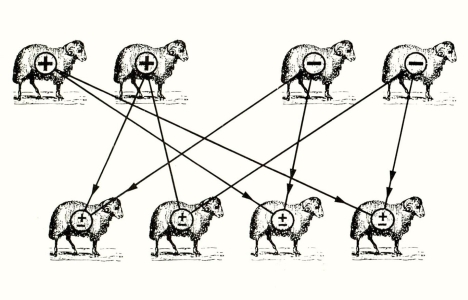
נקח, למשל, עדר כבשים בעלי פרווה דלילה, שאחדים מהם נושאים גנים לפרווה סמיכה (לצורך הדוגמא נניח שמלוא הצופן לפרווה דלילה או סמיכה כלול בגן בודד). מה יקרה, אליבא דמנדל, כאשר איל הנושא גן לפרווה דלילה יזדווג עם כבשה הנושאת גן לפרווה סמיכה? מאחר שכל הורה נושא שני גנים (או אללים) לכל תכונה גופנית, הרי שהסיכוי שיש לכל אחד משני הגנים האלה להמצא בתא המין ההפלואידי (לאחר חלוקת ההפחתה) היא בשעור של 50%. ומכאן, מספר הצרופים האפשריים בין הגנים של האיל לגנים של הכבשה הוא ארבעה.
הדור הראשון של הטלאים שיוולדו לאיל בעל גן הומוזיגוטי (זוג גנים זהים) לפרווה דלילה (+) ולכבשה בעלת גן הומוזיגוטי לפרווה סמיכה (-) יהיו כולם בעלי פרווה דלילה (בתנאי שהגן לפרווה דלילה הוא הדומיננטי), בהתאם לחוקי מנדל (ראה ציור 1). אבל אם נזווג את טלאי הדור הראשון אלו עם אלו, נקבל בדור השני תמונה שונה (ראה ציור 2).
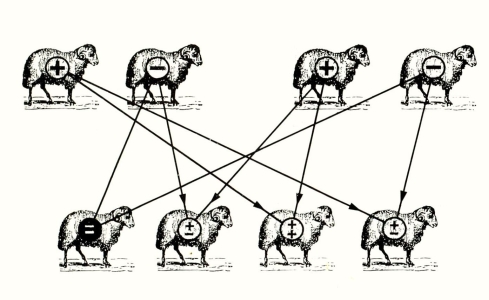
מאחר שטלאי הדור הראשון הם הטרוזיגוטים, כלומר נושאים אללים לפרווה סמיכה ודלילה גם יחד, הרי שבצירוף אחד מתוך הארבעה האפשריים ייצא טלה הומוזיגוטי בעל שני גנים לפרווה סמיכה.
חוקי מנדל מלמדים, איפוא, כיצד יכול טלה בעל פרווה סמיכה להיוולד לשני הורים בעלי פרווה דלילה. מכאן גם ברור כיצד חוקיות זאת, הפועלת על המגוון הגנטי של אוכלוסיה, משרתת את מנגנון הבירור הטבעי. כל עוד תנאי האקלים יציבים תוסיף אוכלוסיית הכבשים בעלי הפרווה הדלילה לפרות ולרבות בהצלחה. אבל אם יחול שינוי אקלימי קיצוני והטמפרטורות יירדו בשיעור ניכר, אזי כף ההצלחה תיטה בצורה ברורה לטובת אותו מיעוט בערר שהוא הומוזיגוטי לפרווה סמיכה. שכן, בגלל עמידותו הגבוהה יותר בטמפרטורה הקרה, יוכל להקים לו צאצאים בעלי כושר השרדות גבוה יותר. עפ״י חוקי מנדל נקבל בדור השלישי התפלגות שונה מאשר בדור השני: 2/3 מהטלאים יהיו בעלי פרווה דלילה, בעוד השליש הנותר יהיה בעל פרווה סמיכה. מכאן, תוך דורות מספר, יפעל מנגנון הבירור הטבעי להפיכת הכבשים בעלות הפרווה הסמיכה לרוב המכריע באוכלוסיה.
הברכה של האנמיה החרמשית
העובדה שאלל רצסיבי יכול לעבור מדור לדור במצב רדום, עד אשר שינוי בתנאים הסביבתיים יעניק לו יתרון הסתגלותי מכריע ויפיץ אותו במהירות באוכלוסיית המין כולו, מקנים לו, מבחינת הבירור הטבעי, מעמד דומה לזה של המוטציה. למשל, כאשר מגדלים אוכלוסיה מסויימת של חידקים בתרבית המכילה סטרפטומיצין, שהוא רעל קטלני מבחינתם, מתקבלת תוך זמן-מה אוכלוסיה עמידה לסטרפטומיצין. עמידות זאת אינה נרכשת עקב כך שהחידקים ״מתרגלים״ בהדרגה לחומר האנטיביוטי, אך גם לא משום שהתחוללה אצלם מוטציה גנטית ההופכת את בעליה למחוסנים מפניו. הסיבה נעוצה באלל רצסיבי בעל תכונת עמידות מפני הסטרפטומיצין, אשר לא הגיע לכלל ביטוי במרוצת מאות ואלפי דורות של צאצאים, עד אשר נקלעו לסביבתו העויינת של הסטרפטומיצין. והנה, די שבקרב אלפי מיליוני החידקים השורצים בתרבית, יימצאו כמה פרטים הומוזיגוטים לעמידות מפני חומר זה, כדי להפוך במהירות את צאצאיהם למרכיבים הבלעדיים של אוכלוסיית חידקים חדשה ומחוסנת מפני הסטרפטומיצין.
אמנם, במרבית המקרים עזרות התכונות הגופניות השונות מפעולה משולבת של מספר גנים, ועם זאת קיימות תכונות לא מעטות וחיוניות ביותר העזרות מפעולתו של גן בודד. כך, למשל, נמצא שנוכחות הומוזיגוטית של הגן ״האפור״ בקרב עכברים, מונעת משיניהם החותכות לצמוח דרך הלסת, דבר המביא למותם כבר בשלב ינקותם. ואילו אצל בני האדם, תופעת ה-RH מינוס בדם עזרת אף היא מצירוף של שני גנים רצסיביים. יכולה כמובן להשאל השאלה, אם אללים אלה הם כה מזיקים, מדוע לא ״דאג״ הבירור הטבעי להכחידם כליל במרוצת האבולוציה? התשובה היא, מפני שהם רצסיבים ורדומים, אך גם מפני שבהרבה מקרים צרוף הטרוזיגוטי זה של אלל דומיננטי (A) עם אלל רצסיבי (A) מקנה לפרט, בנסיבות סביבתיות מסוימות, כושר השרדות גבוה יותר מאשר הצרוף ההומוזיגוטי של שני גנים זהים (AA).8 עדות מענינת ומאלפת לכך היא מחלת האנמיה החרמשית שהזכרנו קודם, שבה כדוריות הרם הפגומות חוסמות את זרימת הדם המקומית וגורמות לנמק של הרקמות. נמצא שליקוי זה של כדוריות הדם מקורו בגן רצסיבי האחראי ליצירת שתיים מתוך ארבע שרשרות ההמוגלובין.
אם כן, כיצר אפשר שגן מסוכן זה לא רק שלא נכחד ע״י הבירור הטבעי, כי אם התפשט בעידודו בקרב 30% מהאוכלוסיה של אפריקה המשוונית? מסתבר שנושאי האלל ההטרוזיגוטי (Aa) עמידים יותר למחלת המלריה, הנמנית עד היום עם המחלות היותר קטלניות באותו אזור, מאשר ההומוזיגוטים (AA) שאינם נושאים את גן האנמיה. אמנם, כפי שמראים חוקי התורשה של מנדל, נשיאת אלל לאנמיה חרמשית (a) פותחת פתח ללידת צאצאים בעלי גן הומוזיגוטי לאנמיה חרמשית (aa), שהיא קטלנית לבעליה – אך לנוכח חומרתה של מחלת המלריה, התוקפת את בעלי הגן AA, פעל הבירור הטבעי להשרדת הצרוף ההטרוזיגוטי Aa בשליש לערך מהאוכלוסיה. המחקרים הראו כי כדוריות הדם של ההטרוזיגוטים, הנושאות בתוכן את טפיל המלריה, מאבדות אשלגן דרך קרום התא ועקב זאת הן מתכווצות ומתקשחות. תהליך זה מביא להריסת דופן התא של הטפילים ומסתיים במותם.

היתרון הגדול של הרבייה המינית
צרופי הגנים המתקבלים, ע״פי חוק מנדל, משיחלוף הגנים בין מערכות הכרומוסומים של ההורים (בעקבות ההפרייה המינית) – מצביעים על היתרון האבולוציוני הגדול הטמון ברבייה המינית, ולפיכך מסבירים את סיבת התפשטותה בקרב הצומח והחי בטבע. שהרי מבחינת פשטות המנגנון וחסכון האנרגיה, ההתרבות הווגטאטיבית (באמצעות חוטרים, למשל) ורביית הבתולים (הצאצאים נולדים מביציות לא מופרות) הן יעילות הרבה יותר.
זאת ועוד, ברבייה א-מינית פטור הפרט מכל הסכנות הכרוכות במציאת בן הזוג, ולא פחות חשוב מזה הצאצאים נושאים את מלוא הגנים של ההורה ולא רק 50% כברבייה המינית. ואמנם, אם הסביבה נהנית מיציבות רבה – יתרונה של רבייה א-מינית ברור. שכן מרגע שהושג ההרכב הגנטי המיטבי, אשר הוכיח את הצלחתו בסביבה נתונה, אין טעם לשנותו. ואכן, מאחר שהצאצאים מהווים כפילים מדוייקים של ההורה, ישמר הרכב זה בכל מהלך הדורות – אלא אם תתחולל בו מוטציה.
לעומת זאת, אם הסביבה נתונה לשינויים ולתמורות, די בשינוי אחד קיצוני כדי להכחיד את האוכלוסיה כולה, באשר היא מורכבת מפרטים זהים ללא גיוון גנטי.
בסביבה לא יציבה יבלוט יתרונה של הרבייה המינית – בראש וראשונה משום ששיחלוף הגנים בין ההורים מבטיח מגוון גנטי עשיר אצל הצאצאים ומאפשר לבירור הטבעי רפרטואר עשיר של אופציות, כפי שראינו בדוגמא של אוכלוסיית הכבשים בעלת האללים לפרווה סמיכה ודלילה.
חשיבותו המיוחדת של שיחלוף הגנים לתהליך האבולוציה באה לביטוי בתנאי סביבה בלתי יציבים, כאשר מנגנון המוטציות האיטי אינו יכול לעמוד בקצב השינויים. יש לזכור כי היחס בין מוטציות מועילות ושליליות הוא 1000:1. כלומר, על כל 1000 מוטציות המזיקות לאורגניזם מתרחשת מוטציה מועילה אחת בלבד. מכאן, מנגנון המוטציות כשלעצמו יתקשה ליצור בסיס גנטי מגוון דיו לפעולה אפקטיבית של הבירור הטבעי והאבולוציה. בסיס כזה מספק שיחלוף הגנים ברבייה המינית.
נניח שבעשר שושלות צאצאים שונות של פרט בודד, המתרבה התרבות וגטאטיבית, ארעו עשר מוטציות באתרים שונים שלאורך הכרומוסום. במקרה כזה יכול הבירור הטבעי לפעול על עשרה גנוטיפים שונים (גנוטיפ – ההרכב הגנטי של פרט מסויים). לעומת זאת, אם עשר שושלות אלו היו מצטלבות ביניהן ע״י רבייה מינית, היו יכולים להיווצר בכל אחר מעשרת האתרים שעברו מוטציה, שלושה גנוטיפים שונים: AA;Aa;aa, ובכל העשרה 310 – שהם 59,049 גנוטיפים אפשריים שונים. במילים אחרות, במקום זירת פעולה מצומצמת של עשרה גנוטיפים, עומדות לבחירת הבירור הטבעי כ-60 אלף אפשרויות התאמה בין האורגניזם לסביבתו.
עכשיו, נמצא שבממוצע שעור ההטרוזיגוטיות (מספר האתרים בכרומוסומים הנושאים גנים לא זהים, קרי: אללים) אצל חסרי חוליות, הוא 13.4%; אצל בעלי חוליות 6.6% ובאדם 6.7%. אם נקבל את ההנחה שקיימים באדם 100,000 אתרי גנים, אזי אנחנו הטרוזיגוטים ב-6,700 אתרים. פירושו של דבר שאנחנו יכולים לייצר בכוח 26,700 (102,017) תאי רבייה שונים!!! לנוכח פוטנציאל אסטרונומי כזה של גיוון גנטי, מובן מדוע אין שני בני אדם (שאינם תאומים) זהים מבחינה גנטית, אבל חשובה הרבה יותר ההוכחה שהוא מספק לתפקיד המרכזי שממלאה הרבייה המינית בתהליך האבולוציה.
בחזרה לאלטרואיזם
כפי שציינו בפתח הדברים, תופעת האלטרואיזם בטבע אינה מתיישבת עם התפישה הדארוויניסטית, לפיה פועל הבירור הטבעי ברמת הפרט – דהיינו כל תכונה המחלישה את כושרו של הפרט להתרבות ולהשרד – נכחדת; ואילו כל תכונה שמועילה לו נשמרת.
היא גם לא מתיישבת עם התפישה הדארוויניסטית9 המודרנית, הגורסת כי תדירות הופעתו של גן מסויים באוכלוסיה תגדל אם גן מסויים זה מגביר את ה״כשירות הדארוויניסטית״ של האורגניזם בכללותו. במילים אחרות: אם יש בו כדי להגדיל בכוח את מספר צאצאיו של הפרט. וכאן בדיוק הקושי. מאחר שרק מי ששורד יכול להנחיל את הגנים שלו לדורות הבאים, כיצד אפשר לו לגן אלטרואיסטי להתפשט ולהתבסס בקרב אוכלוסיה נתונה? שהרי, כל אימת שפעולתו של גן כזה מתממשת, היא מביאה כליה על בעליו.
הבה נניח שלטייסת הקאמיקז היפנית, בשלהי מלחמת העולם השניה, היו נבחרים רק טייסים הנישאים ״גן קאמיקזי״. בתוך שנים ספורות היו נושאי הגן הזה הופכים מיעוט מבוטל באוכלוסיה, הפוחת והולך ככל שהמלחמה נמשכת.
ובכל זאת, למרות המחשה ניצחת זאת, עובדה היא שגנים אלטרואיסטים מסוגים שונים מצאו להם אחיזה איתנה בטבע והם מונחלים מדור לדור זה עשרות מיליוני שנים.
ניקח, לדוגמא, את התנהגותו של כלב הפרא האפריקני, שמינהגי הציד שלו הם שם דבר ל״אכזריות״. כלבי פרא אלה מאורגנים בלהקות שבראשן זכר דומיננטי. לאחר שהמנהיג בוחר בטרף מקרב עדר של גנו, למשל, מסתערים הזכרים לעברו ופותחים במרדף מהיר (עד 65 קמ״ש), הנמשך בין 30 ל-60 דקות. תוך שהם שועטים הם נתלים על קורבנם וקורעים את בשרו עד שהוא מוכרע ומת. והנה, ככל שהציד אכזרי, סיומו אלטרואיסטי להפליא. לאחר שהם ממלאים כרסם בבשר הציד, הם שבים למאורתם ומקיאים את המזון שבקיבתם לפיות הגורים והנקבות ואף חולקים אותו עם זכרים אחרים, שמסיבות שונות לא נטלו חלק בציד. זאת ועוד, בניגוד לאריות, גורי הכלבים נהנים מזכות קדימה בכל הקשור למזון.
נשאלת כמובן השאלה, מה מביא את כלב הפרא האפריקני, בתום ציד מפרך ומייגע, לחלוק את מזונו עם הנקבות, הגורים והבלתי כשירים, ובכך להפחית מכשירותו שלו? ומדוע נוטות הנקבות להניק גם גורים לא שלהן תחת לשמור את מלוא חלבן לצאצאיהן?
לפני שנעיין בתשובה שמציעה הסוציוביולוגיה בסוגיה זאת, הבה נבחן מקרוב דוגמא קיצונית עוד יותר של התנהגות חברתית-אלטרואיסטית, כפי שהיא באה לביטוי בקרב שלוש המשפחות הגדולות מסדרת הדבורניים, סדרה הכוללת את משפחות הנמלים, הצרעות והדבורים.

הדבורה העקרה והמלכה הפוריה
שלוש משפחות אלה מהוות כ-12% מ-8000,000 מינים של חרקים, המקיפים יותר מ-75% מכלל היצורים החיים בעולם.
ייחודן של שלוש משפחות אלה בכך שהן מקיימות (למעט מינים מסויימים של צרעות ודבורים) ״התנהגות חברתית מלאה״, משמע: פרטיהן שומרים על שיתוף פעולה הדוק, פועלים על-פי התמחות מעמדית ומפגינים אלטרואיזם מובהק כלפי המלכה וצאצאיה.
כוורת דבורים עשוייה למנות בין 200,000 ל-800,000 פרטים ומלכה אחת. בהתקרב עונת הנחילים מצמצמת המלכה את הפרשת ה״חומר המלכותי״ לפועלות, דבר המשמש להן, ככל הנראה, אות לבניית מספר מוגבל של תאי מלכות, אשר בכל אחד מהן מטילה המלכה ביצה אחת. משבוקעים מהם הרימות מזינות אותן הפועלות במזון מיוחד המבטיח את התפתחותן למלכות. בפרק זמן זה משתנה מעמדה של המלכה: היא מטילה פחות ביצים, בטנה מצטמקת והתנהגותה הופכת עצבנית. הפועלות, מצידן, מקצות לה פחות מזון ולבסוף אף דוחקות אותה החוצה (במינים מסויימים של נמלים, משמכלה המלכה את חיוניותה מלקקות אותה הפועלות עד שהן מותירות ממנה רק את קליפתה). בעקבות גירושה יוצאת המלכה בראש נחיל של ״נאמנים״, המוצאים להם משכן ארעי כאשכול חי על ענף עץ. ממקום זה נשלחים סיירים לתור אחר משכן קבע לכוורת חדשה.
בינתיים, בכוורת המקורית, בוקעת מתאי המלכות המלכה הבתולה הראשונה, והמעשה הראשון שהיא עושה הוא לבדוק בקפדנות את מצבם של תאי אחיותיה המלכות. כל אימת שבוקעת משם מלכה אחות היא הורגת או מגרשת אותה אל מחוץ לכוורת. משסיימה לבצר את שלטונה הבלעדי, מוציאות אותה הפועלות למסע הכלולות, לעבר הזכרים שממתינים לה בנקודות מפגש מיוחדות. מבחינתו של הזכר המפגש חטוף וקטלני. שכן באקט של ההזדווגות מפוצץ הזכר את מערכת המין שלו בתוך הנרתיק המיני של המלכה ומת לאלתר. המלכה עורכת כשלושה מסעי כלולות ליום, במשך 3-4 ימים. בכל מסע כזה היא מזדווגת עם זכר אחר, ואגב כך צוברת בתוכה כמות זרע שתספיק לה להפרות את כל הביצים שהיא עתידה להטיל במרוצת חייה (כ-1500 ביצים ביום).
אם כן, מכנה חברתי-שתפני זה, שבו המלכה משמשת בתפקיד ״הרחם הקולקטיבי״ של הכוורת, ואילו הפועלות הן הלוחמים, הבנאים, הציידים ומגדלי הצאצאים, צפה ועולה שאלת האלטרואיזם בכל חריפותה. שכן הדבורה המותירה את עוקצה וחלק מכרסה בגוף אויבה, מגינה על הכוורת במחיר חייה. יתירה מזאת, כל חייה היא עמלה בגידול צאצאיה של המלכה – בעוד היא עצמה עקרה. ומכיוון שהיא עקרה, באיזה מובן ניתן להתייחס למעשה ממעשיה כתורם לכשירותה, דהיינו כמעשה המגדיל בכוח את מספר צאצאיה?10 כאמור, מאז דארווין מתחבטים האבולוציוניסטים בסוגיה זאת, אך ללא הצלחה מרובה. והנה, במרוצת עשרים השנים האחרונות התפתחה גישה חדשה, אשר התגבשה לכדי משנה סדורה בסוציוביולוגיה. על פי גישה חדשה זאת יש לחפש את זירת הפעולה של הבירור הטבעי לא ברמת הפרט – על דחפיו, מאבקיו ויצר הקיום שלו – כי אם ברמה קדומה וראשונית הרבה יותר, אשר גבוריה הראשיים הם הגנים האנונימיים.
בירור על-פי שאירים
היו אלה הולדיין (ב-1955) והמילטון (בסדרת מאמרים, 1972-1963), אשר הציעו תשובה לפראדוקס של תופעת האלטרואיזם, דהיינו: כיצד יכול פרט להקריב את חייו למען זולתו, ובכל זאת להגביר בכך את כשירותו שלו?
טען המילטון, מאחר שהגדלת הכשירות נבחנת במספר הצאצאים שהפרט יכול להוליד, ומאחר שמספר הצאצאים מבטאים את כמות הגנים שהוא מותיר אחריו באוכלוסיה נתונה, יחסית לגנים של פרטים אחרים, השאלה אם פרט מסויים הגדיל או הפחית את כשירותו, לא צריכה להמדד ע״י מותו או ע״י מספר הצאצאים שהותיר אחריו, כי אם ע״י שעור הגנים שלו באוכלוסיה בכללותה.
למשל, אם פרט הקריב את חייו כדי להציל אחד מצאצאיו, הוא הפסיד בכך כשירות בשיעור 1 (כל הכרומוסומים שלו) והרוויח 1/2 (50% מהכרומוסומים הנישאים ע״י בנו). במקרה זה עשה עסקה שלומיאלית מבחינת הכשירות, אלא אם-כן הקריב חייו לעת זקנתו – לאחר שאיבד את אונו. אם הקריב חייו למען שני צאצאים, אזי תרם כשירות בשיעור 1 והרוויח 1. במקרה זה מאזן הרווח וההפסד שקול.
לעומת זאת, אם הקריב חייו למען ארבעה צאצאים, אזי תרם כשירות בשיעור 1, אך הרוויח כפול מזה (1/2×4). במקרה זה עשה עסקה מוצלחת מבחינת הגנים שלו. במילים אחרות, מתפישתו של המילטון נובע, כי עקרון פעולתו של הבירור הטבעי – אם לשמר ואם להשמיד – איננו רק מה מועיל ומה מזיק לכשירותו של הפרט, כי אם גם מה מועיל ומה מזיק לכשירות של שאריו – הנושאים חלק זה או אחר של הגנים שלו.
ולפיכך, מבחנה של תופעה או התנהגות גנטית הוא בתרומתה למה שהוא כינה הכשירות הכוללת שלו. הבירור הטבעי הפועל להפצתן או להכחדתן של תכונות – קרוי ״בירור שאירים״.
מהגישה של המילטון נובע, כי התנהגויות אלטרואיסטיות ושתפניות יהיו נפוצות יותר בקרב אוכלוסיות המורכבות משאירים מאשר באוכלוסיות פתוחות בעלות קשרי משפחה רופפים. שהרי בכל מעשה שתעשה למען כלל אוכלוסיית השאירים, אתה תורם במישרין גם למען הגנים שלך הטמונים בפרטים האחרים.
ואמנם, יש תצפיות המאשרות הנחה זו, והיא מקבלת את ביטויה החזק ביותר באוכלוסיות הנמנות עם סדרת הדבורניים, אשר אופן ריבויין מקנה לפרטיהן זהות גנטית גבוהה ביותר.
תאומות של 75%
כזכור מתמלאה מלכת הדבורים בזרע לכל ימי חייה, במהלך מסע הכלולות שלה. הפריית הביצים נעשית תוך כדי מעברן בתעלת ההטלה: מהביצים המופרות יבקעו הפועלות, ואילו מהביצים הלא מופרות יבקעו הזכרים. התוצאה: לזכרים אין אבא, אלא אמא בלבד. מאחר שכך, תאי גופם מכילים רק מערכת כרומוסומים אחת (הפלואידים) במקום השתיים שיש לכל אורגניזם הנולד לשני הורים. פירוש הדבר שכל תאי הזרע הזכריים זהים, באשר כל כרומוסום וכל גן המצוי בכל אחד מתאי גופו של הזכר, מצוי גם בתאי הזרע שלו.
לעומת זאת, תאיה של נקבת הדבורים, הנולדת מביצים מופרות, מכילים שתי מערכות כרומוסומים (דיפלואידים) – אחת מהאב ואחת מהאם. פירוש הדבר שתאי המין של הנקבה זהים רק ב-50%, באשר לכל כרומוסום ולכל גן המצוי בתאי גופן (המכילים שתי מערכות כרומוסומים) יש סיכוי של 50% להיות מיוצג בתא המין (המכיל רק מערכת כרומוסומים אחת).
בגלל הצרוף ההפלואידי-דיפלואידי זה, נוצרים שיעורי קירבה מיוחדים בין הצאצאים להורים ובין הצאצאים לבין עצמם.
שיעור הקירבה (כלומר החלק הממוצע של גנים משותפים) בין הפועלות למלכה הוא 1/2=1/2 1x (הן נושאות אחת משתי מערכות הכרומוסומים שלה). לעומת זאת, שיעור קירבתן כלפי האב הוא 1 = 1×1. דהיינו, בכל הנוגע לאב הן תאומות זהות, באשר הן מקבלות ממנו את מערכת הכרומוסומים היחידה שלו.
הלאה, שעור הקירבה בין הפועלות לאחיהן יהיה רק רבע. מאחר שהזכרים נוצרו רק ממערכת כרומוסומים אחת (מתוך השתים) של המלכה, אזי יחס הקירבה בינם לבין אחיותיהם הוא: מצד האב 0=1×0, ואילו מצד האם 1/2×1/2 (מחצית משתי מערכות הכרומוסומים שלהן זהה עם מחצית משתי מערכות הכרומוסומים של האם). דהיינו: 1/4=(1×0)+(1/2×1/2). והנה, בעוד שיעור הקירבה בין הפועלות לאחיהן הוא רק של 25%, שעור הקירבה בינן לבין אחיותיהן הוא של 75%: הן שותפות מלאות במערכת הכרומוסומים היחידה של האב (1×1/2) ושותפות כדי רבע באחת משתי מערכות הכרומוסומים של האם (1/2×1/2). לפיכך הזהות הגנטית ביניהן היא 3/4=(1×1/2)+(1/2×1/2).1 שעור קירבה גבוה זה אוצר בתוכו, לדידם של הסוציוביולוגים, את המפתח להתנהגותה האלטרואיסטית של הנקבה. לשון אחר, כדאי לה לוותר כליל על פריונה האישי ולהשקיע את מרצה בגידול אחיותיה, הזהות אתה ב-75%, מאשר לפתח מנגנון הולדה משלה היכול להבטיח לה צאצאים נושאי זהות גנטית של 50% בלבד. ומכאן, הקרבת חייה למען שלום הכוורת ושלום מלכתה, היא מעשה נבון והגיוני מצידה; הבא להגן על מאגר הגנים האדיר שלה, המסתכם ב-75% מכלל הגנים של מאות אלפי אחיותיה לכוורת.
אנו רואים איפוא, כי משעה שאנו מעבירים את זירת הפעולה של הבירור הטבעי מהפרט אל הגנים שלו, יכול הלחץ האבולוציוני לפעול לפיתוח מגוון עשיר של תכונות וצורות התנהגות, אשר מבחנן האחד והיחיד נמדד בתועלתן לגנים ולאו דווקא לאורגניזם הנושא אותם.
על כנימות ואריות
כנימת העשבים מהווה דוגמא נאה לכך. בניגוד לחרקים נייחים אחרים, העושים שימוש נפוץ במבני ובצבעי הסוואה כדי לחמוק מחמדנותם של טורפים, תקועות הכנימות בגלוי על העלים וכמו ממתינות באדישות לטורפים שיקטפו אותן. נשאלת, כמובן, השאלה הכיצד לא עודד הבירור הטבעי פיתוח מנגנוני בריחה או הסוואה כלשהם, להגברת כשירותן הדארוויניסטית. התשובה הסוציוביולוגית טמונה, כרגיל, בגנים. הכנימות מתרבות באביב ובקיץ ע״י רביית בתולים (בסתיו – ע״י רבייה מינית) בקצב מדהים. נקבה אחת עשויה, תאורטית, להקים בעונה אחת 600 מיליארד (!) צאצאים המהווים כפילים גנטיים מושלמים שלה (שהרי הם מתפתחים מביצית לא מופרית). או אם תרצה, כנימה שמשקלה מיליגרם אחד יכולה לייצר בעונה אחת צאצאים במשקל 600 טונות. ומאחר שהם העתקים שלה, הרי גם אם נטרפים מיליוני צאצאים, אין זאת אלא אבידה שולית, נגיסה קטנה בגוף האדיר בן 600 מיליארדי פרטים, משל היו מיליונים אלה עלה בודד מעץ עבות. מנקודת מבט של הכשירות הכוללת, גורלם האינדיבידואלי של הפרטים אינו חשוב, אלא כמות הגנים שנשרדת ויכולתם של הפרטים ששרדו להפיץ בתורם הם את הגנים לדורות הבאים.
העברת דגש הפעולה מהפרט לגנים מאפשרת להסביר לא רק התנהגות אלטרואיסטית, כי אם גם את היפוכה. בכך, בעצם, סוד קסמה. למשל, האריות. עדת אריות מורכבת ממספר לביאות בוגרות בעלות קירבה גנטית, אחד עד שלושה זכרים – בדרך כלל אחים, וכמובן גורים.
השתפנות בעדה של אריות עומדת על רמה גבוהה. הציד מתבצע עפ״י חלוקת עבודה מתואמת להפליא והנקבות מתירות גם לגוריהן של נקבות אחרות לינוק מחלבן. בשני אלה התנהגות עדת האריות דומה לזאת של כלבי הפרא האפריקנים. אלא שבניגוד לאחרונים הלביאה ממלאה תפקיד מרכזי בציד, בעוד האריה טורח מעט, אך הוא לעולם ראשון בתור לסעוד. כרגיל, הגורים נאלצים להסתפק בשיירים. אמת, מצבם טוב מעמיתיהם הצבועים, שם חייבת האם לשמור בשבע עיניים שגוריה לא ייטרפו ע״י הבוגרים בבולמוס הסעודה, אך גם על חייהם של גורי האריות שורה תמיד סכנת קטילה. מנהיג העדה רואה בצאצאיו מתחרים מסוכנים, לפיכך הוא מגרשם עוד טרם הגיעם לבגרות מלאה. את בגרותם הם עושים בנדודים – פעם בגפם ופעם בחברת אחיהם – עד שהם מצליחים לגרש או לחסל אריה מבוגר ולהשתלט על עדתו. וכאן קורה דבר מוזר: המנהיג הורג, מעשה ראשון, את כל הגורים של קודמו.11
להתנהגות יוצאת דופן זאת (הרווחת גם אצל קופי הלנגור) מוצאת הסוציוביולוגיה הסבר מעניין: מאחר שתקופת שלטונו של אריה בעדה אינה נמשכת יותר משנתיים-שלוש, יהיה זה מעשה נואל מצידו אם יבזבז זמן קצר ויקר זה בגידול גורים שאינם נושאים את הגנים שלו. יש לזכור שתקופת ההנקה אצל הלביאות נמשכת אף היא 3-2 שנים, וכל עוד הן מיניקות אין הן מתייחמות. הרג גוריהן מאפשר להן איפוא להתעבר מחדש. ואם נוסיף לכך את העובדה שהלביאות ההרות מפילות ולדותיהן בעת המהפך השלטוני, אפשר להבין מדוע ביכר הבירור הטבעי צורה זאת של התנהגות זכרית, שכן היא מגדילה את תפוצת הגנים שלהם. זאת ועוד, הריגת הגורים והפלת הוולדות מבטיחות שהלידה הבאה תהיה מתואמת וכמעט בו-זמנית אצל כל הנקבות. משמע, טיפול טוב יותר לגורים החדשים.
שאלת הזהות הגנטית של הצאצאים היא אחד המרכיבים המרכזיים בעיצוב דפוסי ההתנהגות של בעלי חיים. הבירור הטבעי, מסתבר, אינו מעודד השחתת אנרגיה והוא בוחן כל התנהגות על-פי תרומתה להגברת הכשירות הכוללת של בעל הגנים. מסיבה זאת תופעות של ״בעלים קרננים״, או מעשי הונאה דוגמת הקוקיה (המטילה ביציה בקינים זרים), הינם נדירים למדי. שכן, כל יחידת אנרגיה שהאורגניזם משקיע בגידול צאצאים לא לו, פירושה פחות יחידת אנרגיה (ובזבוז יחידת זמן) בגידול צאצאים הנושאים את הגנים שלו.
הסיבה למגמה זאת של הבירור הטבעי כמעט ואינה דורשת הסבר. נניח שהיו בעבר אריות שלא נשאו צו גנטי לחיסול גורי קודמיהם, אלא נהגו כהורים מאמצים לכל דבר. התוצאה, בתום ״כהונתם״ כראשי העדה הותירו אחריהם אך צאצאים ספורים. צאצאים אלה, בתורם, אמצו אף הם גורים לא להם, בהגיע תורם להשתלט על עדות חדשות, וכך חוזר חלילה. ניתן להניח כי בתום מספר דורות נעלמו כליל נושאי הגנים ״ההומניסטים״ האלה מקהילת האריות.
עקרון זה של כדאיות ההשקעה עומד ביסוד המגוון העצום של התנהגויות בעלי-חיים בטבע.

על שוביניזם זכרי וחגורות צניעות
יוני הטבעת מגלים רגישות שוביניסטית מופלגת בכל הקשור לצניעותן של כלותיהם לעתיד. הסיבה לכך נעוצה בניגוד האינטרסים בין הזכר לנקבה. מנקודת מבטה של הנקבה, מרגע שהופרתה, אין נפקא מינה אם בן הזוג שיבנה עמה את הקן וידאג למחסורה בעת הדגירה הוא הזכר שהפרה אותה או כל זכר אחר, שהרי בכל מקרה הגנים שלה מובטחים בביצים שהיא עתידה להטיל. לא כן הזכר. לגבי דידו, שאלת תומתה של בת זוגו היא קרדינלית. שכן, אם כבר הופרתה ע״י זכר אחר, כל המאמץ שהוא השקיע ועתיד להשקיע בחיזור, בבניית הקן, בהגנה עליו ובגידול הגוזלים, יהיה מבחינתו לריק ורק יתרום להפצת הגנים של זכר אחר.
הטחביות (סרטן יבשתי שחי מתחת לסלעים), פתרו את הבעיה בשיטה פשוטה. הזכר משתלט על הנקבה מספר ימים לפני ההפריה, לופת אותה בין רגליו הרבות ונושא אותה צמודה אליו לכל אשר ילך. בכך הוא מבטיח לעצמו, כמובן, זכות הזדווגות בלבדית. אצל יוני הטבעת נועד טכס החיזור לבחון את סוגיית הנאמנות של בת הזוג: האם היא תמה או נפקנית? טכס החיזור של הזכר גורם לנקבה להפריש סטרואידים, שמביאים אותה להציג תנועות טכסיות של בניית קן. אם הוא מתמיד בחיזורו מסתיים העניין בהזדווגות. והנה, בניסוי שערכו, הפרידו את הנקבות מהזכרים לאחר השלב הראשוני, ולאחר זמן-מה הפגישו אותן עם קבוצה אחרת של זכרים. הנקבות, כמובן, הפגינו נכונות להזדווג עימם לאלתר, שהרי כבר היו למודות חיזור מקודם. אלא שנכונות זאת היתה חשודה בעיני הזכרים, באשר העידה על עברן ״המפוקפק״ ועל האפשרות שהן כבר נושאות בתוכן זרע זר. לפיכך מאנו הזכרים ברוב המקרים להזדווג עמן, לפי הכלל העממי: מה שבטוח בטוח.
מינים רבים של נחשים, תולעים ואף עכברושים וחזירוני-ים נוקטים בשיטה ישירה ובוטה יותר להבטחת צניעותן של בנות זוגם: בהמשך להפרייה הם מפרישים חומר מיוחד המתקרש ויוצר פקק בפתח המיני של הנקבה ומונע מזכר אחר להפרותה. מינים אחרים אף הגדילו לעשות: הזכרים תוקפים אלה את אלה והחזקים יותר מחדירים במנוצחים חומר פיקוק המונע מהם הפרשת זרע. אצל מינים מסויימים של זבובים הזכר מחדיר את זרעו בזכר אחר – בעת שהלה מפרה נקבה – ומשתמש בגופו כבצנור לזרעו שלו. בקרב השפירית האמריקנית משתמש הזכר באבר המין שלו דמוי יעה בקצהו, כדי לגרוף מתוך גופה של הנקבה את הזרע של קודמיו, בטרם ישאיר את שלו למשמרת. אצל עכברים, נקבה מעוברת שמשתכנת במחילה עם עכבר אחר, מפילה את עובריה בהשפעת הריח המיוחד שמפריש הזכר, ובכך הופכת כשירה להתעבר ממנו. תופעה מדהימה הרבה יותר מתרחשת אצל היבחושים העוקצניים. בעקבות ההזדווגות טורפת הנקבה את הזכר, למעט את אבר המין שלו, הנותר תקוע בפתח הנקבי שלה, דבר המונע מזכר אחר להזדווג עימה. ואם כבר אנו עוסקים בקניבליזם מצד הנקבה ובנכונותו ההרואית של הזכר להקריב חייו על מזבח הגנים שלו (כנהוג בקרב מינים של עכבישים), אי אפשר שלא להזכיר את פרשת היחסים המאלפת בין הזכרים והנקבות של גמלי שלמה. בדרך כלל מחסלת הנקבה את הזכר תיכף להזדווגות ונהנית בכך לא רק מהפרייה, כי אם גם מארוחה דשנה ועשירת חלבונים. אך לעתים רוחה קצרה והיא מולקת את ראשו של הזכר עוד טרם ההפרייה. והנה, הפלא ופלא, לא רק שעריפת הראש אינה מפריעה לזכר למלא את תפקודו המיני, כי אם גם משחררת אותו מהפחדים ומההיסוסים המאפיינים את גישתו לנקבה. אם תרצה, דרך יעילה, אף כי דרסטית, למניעת אימפוטנציה בקרב הזכרים.
ואולי נסיים מדגם מצומצם זה מתוך הרפרטואר המיני העשיר והמגוון להפליא של הטבע, בסיפורו של הברכיה (סוג ברווז). הברכיה מקיים קשר מונוגמי לכל החיים עם בת זוגו, קשר רצוף גילויים מופלאים של נאמנות ומסירות. הוא גם נלחם בשצף-קצף עם זכרים המנסים להפרות את בת זוגו, אך באותם מקרים שידו על התחתונה הוא מצטרף לתוקפים, במה שאפשר לכנותו אונס קבוצתי, כדי לתת סיכוי כלשהו גם לזרע שלו.
זכר לנקבה – זאב
מגוון ההתנהגויות המיניות ומערכות היחסים בין הזכרים לנקבות, כגון זאת הקיימת בין יוני הטבעת, מצביעה על אחת הסוגיות המרתקות ביחסי המינים: ניגודי האינטרסים בין הזכר לנקבה, שראשיתם ברמת הביצית ותא הזרע.12 אם מטרת היסוד של כל אורגניזם היא להפיץ כמות מירבית של גנים שלו, חייב להיווצר קונפליקט משמעותי – ולפיכך גם שוני בהתנהגות – בין הזכר לנקבה, שורשו של הקונפליקט הזה מתגלה כבר בגודל ובתפקוד של תאי המין. הזכר נושא כמות אדירה של תאי זרע זעירים, אשר לייצורם השקיע אנרגיה מעטה יחסית. ומאחר שהוא פטור מנטל נשיאת העובר בתוכו, האסטרטגיה שתשרת בצורה מירבית את מטרת היסוד שלו – היא להפרות מספר גדול ככל האפשר של נקבות ולהסתלק מהמקום תיכף להזדווגות, פן יאלץ לחלוק עם הנקבה את נטל הדאגה והגידול של צאצאיהם המשותפים. במילים אחרות, הלחץ האבולוציוני היה צריך להפוך את כל הזכרים בטבע לדון ז׳ואנים, השוהים במחיצת הנקבה לא יותר מן הזמן הדרוש להפרותה, וממשיכים מיד הלאה לנקבה הבאה בתור.

מטרת היסוד של הנקבה זהה לזאת של הזכר: להפיץ כמות מירבית של גנים שלה, אך מצבה שונה בתכלית השוני משלו. ראשית, היא משקיעה בייצור הביצית הגדולה הרבה יותר אנרגיה מאשר נדרשת לזכר לייצר את זרעוניו; שנית, משהופרתה אין היא יכולה להתעבר פעם נוספת אלא רק בתום ההטלה או הלידה; שלישית, כמי שנושאת את העובר בבטנה היא חייבת בהזנתו.
על פי עקרון הכדאיות שהזכרנו קודם, לא יהיה זה נבון מצידה לנטוש את הוולדות לאחר שהשקיעה בהם אנרגיה כה רבה. (לפי גרסה אחרת, הנטישה בשלב זה אינה כדאית לה, כי ההשקעה הנדרשת לשמור על הקיים ולהביא למימושו, קטנה יותר מההשקעה שתידרש לה כדי לחזור ולהגיע למצבה הנוכחי). מכאן, האסטרטגיה מצידה תהיה לאלץ את הזכר הדון-דואן להשקיע זמן ומאמץ להשגתה – אם ע״י טכסי חיזור מייגעים ואם במאבקים מתישים ביריביו הזכרים – כך שעניינו בטובת צאצאיו, לאחר כל האנרגיה שהוציא לצורך הפרייתה, יהיה לא פחות משלה. ואמנם, העוקב אחר המאמצים והזמן שמשקיע הצפור הזכר בחיזור אחר הנקבה ובבניית הקן, ברי לו שחשבון הרווח וההפסד של השקעתו יורה לו בעליל כי כדאי לו להמשיך לדאוג למחסורה ואף להשתתף בהזנת הגוזלים, מאשר להתחיל את כל הסיפור המייגע הזה עם נקבה חדשה ועוד להסתכן באבדן צאצאיו הנושאים את הגנים שלו.

אבל, אם הארכנו בתאור התנהגותו הנלוזה של הזכר בטבע, מן הדין להביא דוגמא אחת של התנהגות נלוזה מצד הנקבה. הז׳אקנה האמריקנית היא עוף מים המקיימת קשר פוליאנדרי (מקביל לקשר הפוליגמי אצל הזכר) עם מספר זכרים. בהגיע עונת הקינון היא משתתפת עם הזכר בבניית הקן, אבל רק לכאורה. למעשה אין היא תורמת דבר פרט לתנועות-סרק ומעט סכך. לאחר שהיא מטילה ביצים בעבור הזכרים שלה, היא משאירה להם את מלאכת הדגירה הנמשכת 28 ימים.
כאמור, התנהגותה של הז׳אקנה, וככלל – תופעת הפוליאנדריות בטבע, היא בגדר חריג המלמד על הכלל. והכלל הוא שכושר הפרייתו של הזכר אינו מוגבל אלא ע״י אונו וזריזותו, בעוד עניינה של הנקבה בזכרים מסתיימת ברגע שהופרתה.
אולם, קביעה זאת אינה יותר מאשר צילום המצב העובדתי בשטח ואין היא מסבירה את סיבת היווצרותו של מצב המקנה כבר ברמת תאי הרבייה יתרון ניכר לזכר. שהרי ניתן לשאול: מאחר שאת מיזוג הכרומוסומים של הזכר והנקבה אפשר להשיג גם ע״י ריבוי איזוגמי (רבייה מינית שבה הפרטים אינם נבדלים לזכרים ולנקבות), מדוע אין הביציות של הנקבה זעירות וניידות כמו תאי הזרע של הזכר?
לדברי טים הולידיי לא התפתחו הדברים כך משתי סיבות: במאבק הקיום החריף המלווה את האבולוציה של תאי הרבייה, פעל הבירור הטבעי לטובת שתי תכונות:
א. זו שהגדילה את סיכויי התא למצוא את בן זוגו.
ב. זו שהאריכה את תוחלת חייו של התא.13
במילים אחרות, הבירור הטבעי עודד התפתחותם של תאים בעלי ניידות גבוהה ואריכות ימים. אולם, מאחר שהתכונה הראשונה דורשת גוף זעיר ואיבר תנועה, ואילו התכונה השניה דורשת גוף גדול בעל אספקה עצמית מבחינת עתודות המזון, לא היה אפשר לשלבם בגוף אחד, אלא למיינם בשני גופים נפרדים. התוצאה: הנקבה מביאה לזיווג מעון נוח ונדוניה נכבדה של מזון, בעוד הזכר תורם, כפרזיט מושלם, רק זרע ארעי אך זריז ונייד.

הקשר האכזרי והדאגה האמהית
ניגוד אינטרסים דומה שורר גם בין הורים לצאצאים, למרות הביטויים האלטרואיסטיים המספקים השראה למשוררים. הצאצא הוא נושא הגנים של ההורה, אך בו-בזמן גם יריבו למזון ולהגמוניה על הנקבות; ההורה מחויב גנטית לדאוג ולטפל בצאצאיו, אך בו בזמן הם בעבורו גם מזון עשיר וקל-טרף. למעשה מתעורר הקונפליקט תיכף ללידה: האינטרס של הצאצא הוא להאריך ככל האפשר את תקופת היניקה, בעוד האינטרס של האם הוא לגמול אותו בהקדם, הן משום שהדבר מטיל נטל כבד על משאביה והן כדי לשחררה להקמת דור חדש של צאצאים.
ב-1972 טבע החוקר טריברס את המונח ״השקעת הורים״, שמשמעה כל השקעה של הורים בצאצא, המגדילה את סיכויי השרדותו על-חשבון יכולתם להשקיע בהקמת צאצאים נוספים. לפי המודל של טריברס כדאי לאם לטפל בצאצא שלה, כדי להבטיח את השקעתה בו, עד לאותה נקודה שיוכל לדאוג לעצמו. מנקודה זו ואילך כדאי לה להקדיש את מרצה ואונה להקמת דור חדש של נושאי גנים שלה. מעבר זה בין קוטבי הכדאיות מתממש במינים שונים באופן הדרגתי. למשל, אצל כלבים וחתולים, בשלב חייהם הראשון, יוזמת האם את ההנקה, ואילו בשלב מאוחר יותר עוברת היוזמה לידי הגורים; בשלב הגמילה תובעים הצאצאים במפגיע את היניקה ואילו האם דוחה אותם מעליה בחריפות גדלה והולכת. לדעת החוקר הישראלי אמוץ זהבי, מקבל קונפליקט זה אופי של ״סחיטה שטנית״: הצאצא תובע את מזונות בקולי קולות כדי למשוך את תשומת ליבם של טורפים שכנים, ותחת איום זה נאלצת האם להשביע את רעבונו. זהבי גורס, כי הבירור הטבעי מעודד את התפתחותם של גנים ״סחטנים״ כאלה, שכן צאצאים הנושאים אותם נהנים מתזונה משופרת ולפיכך משיגים ״כשירות דארוויניסטית״ גבוהה יותר. טריברס, מצידו, עומד על ביטוי אחר של יחסי הורים-צאצאים, אותו הוא מכנה ״הקשר האכזרי״, הנובע מהקונפליקט המסורתי בין זכרים לנקבות.
כאמור, האינטרס הבסיסי של הזכר הוא להסתלק תיכף להפריית הנקבה; האינטרס של הנקבה הוא להסתלק מייד לאחר הלידה/הטלה. הטקטיקה הטובה ביותר היא לנטוש ראשון, שכן המקדים לנטוש אינו מסכן את עתיד צאצאיו (דהיינו – השקעתו), אלא רק מותיר את האחריות עליהם לבן זוגו. לנטוש שני פירושו לחרוץ דין מוות על הצאצאים (דהיינו אבדן הגנים שלו). הזכר, כאמור, יכול להסתלק מייד בתום ההזדווגות ולתור לו אחר נקבה חדשה להפצת מחזור נוסף של גנים שלו. לא כן הנקבה. שהרי בשל ההריון והלידה היא ״האחרונה בשטח״ ולפיכך קשורה לצאצאיה ב״קשר אכזרי״: אם תנטוש אותם תיכף ללידה – תאבד את השקעתה; אם תישאר – תיאלץ להשקיע את כל אונה ומרצה לגידולם, במקום להתפנות לזכר חדש וללידה נוספת.
יש הרואים ב״קשר האכזרי״ את הבסיס הביולוגי למה שמכונה בפינו ״הדאגה האמהית״, אשר הטבע כה מלא בגילוייה.
הכי ניתן לאמת תיזה זו? אולי. אם נגלה תופעה הפוכה של ״דאגה אבהית״, אצל זכרים הקשורים לצאצאיהם ב״קשר אכזרי״. והנה, מסתבר שתופעה כזאת קיימת בקרב מינים רבים החיים בים. בניגוד לחיים ביבשה, במים לא נשקפת סכנת התייבשות לתאי הזרע. לפיכך יכול הזכר להפרות את ביצי הנקבה מחוץ לגופה, לאחר שהיא מטילה אותן במים. מאחר שכך הנסיבות פועלות לטובת הנקבה הימית: היא זאת היכולה לממש את האינטרס היסודי שלה ולהסתלק תיכף להטלת הביצים: ואילו הזכר נותר ״אחרון בשטח״ ולפיכך הוא נושא לבדו במלוא האחריות לחובת ההפרייה. לכאורה, יכול הוא להקדים ולשחרר את זרעו באזור ההטלה המשוער של הנקבה, אך מאחר שהזרעונים קלים יותר מהביצים – עלולים הם להתפזר במים בלי למלא את יעודם. התוצאה: נטל ״הקשר האכזרי״ נופל עליו. ואמנם, בקרב מינים רבים של דגים רווחת יותר ״הדאגה האבהית״ מאשר ״הדאגה האמהית״. יתירה מזאת, במקרים רבים עליו להתחכם לנקבה כדי להבטיח את מעשה ההפרייה. לדוגמא, במינים מסויימים של דגים, כמו האמנונית, מדגירות הנקבות את הביצים בלוען. כיוון שכך הן נוהגות לאסוף כל גוף כדורי לבן המצוי בקרבתן – שמא זאת ביצה שנשמטה מפיהן. הזכרים ״למדו״ במרוצת האבולוציה לנצל נטייה זאת. הם פיתחו בסנפיר האנלי שלהם כתמים לבנים עגולים, ובהתקרב הנקבה לעברם הם מציגים אותם לראווה. היא, בדמותה שכתמים לבנים אלה הם ביצים נטושות, שוחה לעברם כדי לאסוף אותן בפיה. זהו הרגע לו חיכו הזכרים: הם מתיזים לעבר פיה הפעור רסס זרע המפרה את הביצים הטמונות בלועה. אגב, צורת הדגרה עוד יותר מוזרה מאשר הלוע היא השימוש שעושה צפרדע אוסטרלית זעירה (55 מ״מ) בקיבתה. קרפדה זאת בולעת את הביצים המופרות והופכת את קיבתה לרחם זמני. הראשנים בוקעים מהביצים ומתפתחים לצפרדעים זעירות בקיבתה. בטנה של האם מתפתחת למימדים כה אדירים עד כי ריאותיה מתמוטטות והיא נאלצת לנשום דרך עורה. ואז, בהגיע המועד, נורים עשרות צאצאים מפיה כמתוך לוע של תותח, בקצב של 4 צפרדעים בשניה ולמרחק של עד 60 ס״מ (!).
הדילמה של קוף הגלאדה
מצבי קונפליקט וניגוד אינטרסים, הניצבים בפני בעלי-חיים על כל צעד ושעל, מספקים את אחד הפרקים המרתקים והמאלפים בספר הטבע.
באבונים ממין הגלאדה חיים בקבוצות המונות מנהיג אחד וכעשרים נקבות וצאצאים. האינטרס של המנהיג הוא להרבות בנקבות כרי להבטיח תפוצה מירבית לגנים שלו. אלא שאליה וקוץ בה. הנקבות נותרות14 עימו כל עוד הן זוכות ממנו למנה ראויה של תשומת לב. משהן חשות עצמן זנוחות הן חדלות לפלות כינים אחת לרעותה, אחדות הקבוצה מתפוררת והן נוטות אז להעתר לחיזורי הזכרים החופשיים המשוטטים בקרבת מקום.
וכאן הדילמה: ככל שמספר הנקבות שברשותו גדול יותר, חייב המנהיג להקדיש (בדרן כלל) יותר זמן למלחמה בזכרים הזרים: מצר שני, ככל שהוא עסוק יותר במלחמות – נבצר ממנו להקדיש להן תשומת לב. התוצאה – הזכר נקלע למצוקה קשה מנשוא: הוא אץ לכל עבר במאמץ לגרש את הזכרים הפולשים, ובו בעת מתרוצץ כל עוד נפשו בו מנקבה לנקבה כדי להפיס דעתן.
זיווגים מתימטיים
אחת הדילמות הקשות הניצבות לפני זכרים רבים בטבע נוגעת להשקעה היחסית של הורים: במה כדאי להשקיע יותר זמן ואנרגיה – בהבטחת ההפרייה ובגידול הצאצאים העתידים להנחיל את הגנים שלך לדורות הבאים, או בהפריית מספר גדול ככל האפשר של נקבות (בהנחה שדי גם בשרידות נמוכה של צאצאים כדי להבטיח את ההמשכיות שלך)?
ד״ר גאופרי פרקר בחן שאלה זאת בקרב זבובי הזבל, המקיימים את פעילותם המינית סביב צואת בקר טרייה. משמאתר הזכר נקבה פנוייה הוא נוחת על גבה ומזדווג עימה קצרות. לכאורה יכול הזכר להזדווג בשיטה זאת עם מספר עצום של נקבות. אלא שאין הדברים פשוטים כל כך. מאחר שהביצים מופרות תוך כדי מעברן בצנור ההטלה של הנקבה (בדומה להפרייה אצל מלכת הדבורים), הרי שאחרון להחדיר זרעו בצנור הוא גם זה שמזריע כדי 80% מהביצים של הנקבה. הזכר ניצב איפוא לפני דילמה קלאסית: אם ידבוק בנקבה למשך שלוש שעות (בקירוב) החולפות בין ההזדווגות להטלה, יבטיח אמנם את הפריית הביצים בזרעוניו, אך יחמיץ בכך עשרות נקבות אחרות ויאבד את הסיכוי להשכין את הגנים שלו במאות אלפי ביצים.
ובכן, מה תהיה האיסטרטגיה המיטבית בשבילו? במילים אחרות, כמה זמן כדאי לו לשהות על גב הנקבה, כדי למנוע אותה מזכרים אחרים, ועם זאת להזדווג עם מספר מירבי של נקבות? ד״ר פרקר וצוותו ניסחו את השאלה בצורה מתימטית במסגרת תורת המשחקים.2
התשובה הורתה על 41 דקות כעל משך הזמן האופטימלי שעל הזכר לשהות על גב הנקבה – אם ברצונו להפיק את המירב משני העולמות. והנה, כאשר בדקו את המצב בשטח ומדדו את זמן השהייה הממוצע של הזכר על גב הנקבה, מצאו שהוא 36 דקות. 4 דקות פחות מהזמן המיטבי שהתקבל במודל המתימטי.
יתירה מזאת, החישובים המתימטיים הראו שהזדווגות עם נקבה שזה עתה הטילה כדאית פחות מהזדווגות עם נקבה שכרסה מלאה ביצים. (במקרה הראשון גדול יותר הסיכוי שעד למועד ההטלה היא תזדווג עם זכרים נוספים). לפיכך כדאי לזכר לשהות רק 18 דקות על גבה של נקבה משוללת ביצים, לעומת 36 דקות על גבה של נקבה גדושת כרס. ואמנם, כאשר נבדקה המציאות בשטח, התברר שהזכרים אכן שוהים רק 15 דקות במחיצתן של נקבות ריקות כרס.
האם מסקנת המחקר הזה היא שזבובי הזבל מחשבים חישובים מתימטיים בראשם? כמובן לא. הממצאים עשויים להצביע על כך שהבירור הטבעי מבכר תכונות והתנהגויות הנותנות תוצאות חיוביות15 מבחינת ״הכשירות הדארוויניסטית הכוללת״ של בעליהן. העובדה שהאופטימיזציה פועלת עפ״י כללים מתימטיים מסבירה את התאימות הגבוהה בין הניבוי של המודל המתימטי לבין המציאות בפועל.
מאחר שכך ניתן להציג שאלה נוספת: האם גם התנהגויות אחרות, המוכתבות מבחינה גנטית, פועלות על-פי כללים מתימטיים?

גנים יוניים וניציים
אין זאת שכך נראו הדברים למיינרד סמית בהטביעו את המונח Evolutionary Stable Strategy (ESS), דהיינו: שיווי משקל תחרותי (המהווה גרסה של תורת המשחקים). מטרתו של מיינרד סמית היתה לבנות מודל מתימטי שיצביע, במצבי תחרות בין חיות, על אסטרטגיית פעולה (ESS), שאם תאומץ ע״י מרבית החיות באוכלוסיה נתונה תקנה להן כשירות דארוויניסטית גבוהה יותר מכל אסטרטגיית פעולה אחרת, או מכל שינוי ותיקון שיכניסו באסטרטגיית הפעולה של ה-ESS.
ואם אנו מאמצים ההנחה שהבירור הטבעי מעודד אופטימיזציה של התנהגויות, פירוש הדבר שה-ESS שנקבל מהמודל המתימטי יאפשר לנו לנבא באילו דרכי פעולה ינקטו בעלי החיים בתסריטים שונים של המציאות הטבעית. יתירה מזאת, הוא יאפשר לנו להבין מדוע קיימים בטבע דפוסי התנהגות כאלה ולא אחרים. למשל, מרבית המאבקים התוך-מיניים בטבע אינם מגיעים לכלל עימות פיסי, הם מוכרעים באמצעות טכסי-כוח וראווה שונים, שמטרתם להטיל מורך בלב היריב ולגרום לו להסתלק.
ניתן, כמובן, לשאול: מה מונע את אחד המתמודדים להעמיד פני נכנע ולתקוף את יריבו כאשר זה נפנה ממנו? במילים אחרות – מדוע אין עושים שימוש בהתמודדויות אלה במעשי הונאה? ואם אמרנו שהאלטרואיזם משתייר בטבע הודות לתרומתו לכשירות הכוללת של הפרט, מרוע לא ישתייר גם ״גן נוכלי״ שיש בו כדי לתרום לכשירותו של הפרט ע״י מעשי הונאה?
תשובתו של מיינרד סמית: ESS.
להבהרת רעיונו הציג סמית את הדוגמא הבאה: הבה נכנה בתואר נץ, פרט שנוטה להסלים כל עימות שהוא נקלע אליו – עד לנצחונו או לפציעתו. ואילו בתואר יונה נכנה פרט הנוטה לברוח כל אימת שיריבו מסלים את המאבק. עכשיו, ברור כי אסטרטגיה ניצית תעשה חיל באוכלוסיה של יונים, אך היא תניב פירות באושים באוכלוסיה ניצית. שהרי תוך זמן קצר יסתבכו הניצים במאבקים ״עד הסוף המר״ וייצאו מהם בשן ועין. קיימת, כמובן, אסטרטגיה שלישית, שהיא תערובת של יוניות וניציות, דהיינו: אם יריבך קטן ממך – תקוף; אם הוא גדול ממך – ברח; אם הוא במידותיך – פעל בהתאם לנסיבות (אם הוא מסכן את גוזליך וכיו״ב).
אם-כן, מהי אסטרטגיית הפעולה אשר תאומץ ע״י אוכלוסיה, בגין הכשירות הדארוויניסטית המיטבית שהיא מבטיחה לכל הפרטים? הבה נראה את הדברים במודל מתימטי.
נתחיל בכך שכל עימות משפיע על הכשירות הדארוויניסטית של המתמודדים. ניתן איפוא לתת ערך מספרי להשפעות אלה. לדוגמא:
10+ נקודות: נצחון 20- נקודות: פציעה
3- נקודות: בזבוז אנרגיה וזמן (כמו בעימות בין יונים המסתובבים אחד סביב רעהו ללא הסלמה).
אם-כן, בעימות בין שני יונים, יבזבזו השניים זמן ואנרגיה, אך אף אחד מהם לא יפצע, ועדיין יש לשני היריבים הסתברות של 50% לנצח. ובכן, התמורה הצפוייה ליונה באוכלוסיה המורכבת מיונים בלבד, היא:2+=(3-)+(10+)(1/2).
עכשיו, אם באוכלוסיה של יונים תיפול מוטציה באחד הגוזלים, שתקנה לו תכונה ניצית, הוא יהנה מיתרון עצום, שכן בכל מפגש עם עמיתיו היונים ייצא ברווח של 10+. הואיל וכך, צאצאיו, אשר יירשו את הגנים הניציים, ייהנו אף הם מכשירות דארוויניסטית גבוהה ויצליחו לפיכך להפיץ הגנים הללו במהירות רבה.
אבל מה יקרה כאשר האוכלוסיה כולה תישא בקרבה גנים ניציים? בכל עימות תהיה לכל אחד מן המתמודדים הסתברות של 50% לנצח – אך גם סיכוי של 50% להפצע, סיכויי הרווח הממוצע יהיו איפוא:5-=(20-)(1/2)+(10+)(1/2)
נהפוך עכשיו את היוצרות ונעבור לאוכלוסיה ניצית. מה יהיה גורלו של גוזל שיוולד עם מוטציה יונית?
הוא לא ינצח לעולם – אך גם לא יפצע (לפי שיתחמק מכל עימות). לפיכך הרווח שלו יהיה אפס. אבל רווח אפס עדיף על רווח של 5-, שהוא מנת חלקם של עמיתיו הניצים. הואיל וכך, הגן היוני יתפשט עד מהרה באוכלוסיה הניצית, עד שתיהפך ברובה יונית, דבר שיקנה יתרון של 10 + לגן הניצי, וכך חוזר חלילה: אוכלוסיית בעלי הכנף תעבור הלוך ושוב מיוניות לניציות בתנועה מתמדת של מטוטלת.
מסקנה: לא התנהגות ניצית ולא התנהגות יונית יכולות להוות בסיס לאסטרטגיה יציבה (ESS) , אשר תאומץ ע״י האוכלוסיה בכללותה.
מהו, אם כן, ה-ESS לעימותים בין יונים לניצים? אסטרטגיה מעורבת של יוניות וניציות במינון מסויים: על כל 13 מפגשי עימות – נהג כנץ 8 פעמים וכיונה 5 פעמים.
כמובן, האסטרטגיות הנקוטות בטבע מורכבות יותר מאשר קווי פעולה פשטניים נוסח ״נהג כנץ״ או ״נהג כיונה״, או אפילו תערובת של השניים. למשל, בעימות בין שני ״יונים״ על מקור מזון יכול אחד מהם לגלות ניציות יתירה עקב היותו רעב יותר מיריבו, או משום היותו בעל המזון השנוי במחלוקת. התנהגות אחרת, הרווחת מאוד בטבע, באה לביטוי במאבקים בין בעלי טריטוריות לפולשים הזרים. קו הפעולה המאפיין עימותים אלה הוא כלהלן: ככל שהפולש חודר עמוק יותר ללב הטריטוריה של יריבו, הופך האחרון ניצי יותר וידו גוברת. ולהיפך – פולש ניצי יהפוך יוני יותר ככל שהוא מתרחק מהטריטוריה שלו.
דוגמא מעניינת ומקסימה ליתרון ההתמודדותי שמקנה הבעלות הטריטוריאלית, ניתן למצוא בקרב פרפרי החורש המוכתמים, שמקום מושבם באירופה. פרפרים אלה חיים בחורשות עבותות המאפשרות רק לקרני שמש בודדות לחדור בעדן ולשבץ את רצפתן המוצלת בכתמי אור. מסתבר שכתמי אור אלה משמשים אתרי חיזור מועדפים, ומאחר שמספרם קטן תמיד ממספר הזכרים, נאבקים הללו ביניהם על הבעלות על כתמי האור. כל אימת שפרפר זר פולש לכתם אור, יוצא בעליו נגדו והשניים מתעופפים בתנועה לוליינית לעבר צמרות העצים. אך תוצאות המאבק קבועות מראש: לעולם מנצח הבעלים של כתם האור. אלא שאין זאת בעלות קבע. די שהבעלים יעדר מכתמו לדקות ספורות כדי שיאבד את חזקתו. לכשינסה לחזור לאתרו – ינוצח ע״י הבעלים החדש.
אם רוצים, אפשר למצוא במודל ה-ESS הסבר גם לחלוקה השווה השוררת בטבע בין זכרים לנקבות, שאינה מתיישבת, לכאורה, עם נטייתו ויכולתו של הזכר להפרות מספר גדול של נקבות. אצל כלבי-ים, למשל, 88% מהנקבות בעדר מופרות ע״י 4% בלבד מהזכרים. מכאן, האינטרס וחשבון הכדאיות צריכים היו להביא את ההורים לשאוף להוליד מספר גדול ככל האפשר של זכרים על חשבון נקבות. החשבון פשוט: כל השקעה בבת תכניס לקופת ההורים שכפול אחד של מערכות הגנים שלהם באמצעות יוצא חלציה. לעומת זאת, אם ישקיעו בבן, הוא עשוי להמנות על 4% המובחרים שיפרו 88% מנקבות העדר. ואם נניח שהעדר מונה 100 נקבות, פירוש הדבר שבן יקיר כזה עשוי להכניס לקופת ההורים 22 שכפולים.
מדוע, למרות זאת, מורכבת אוכלוסיית כלבי-הים שווה בשווה מנקבות ומזכרים? משום שמתן עדיפות להולדת זכרים אינה יכולה להוות אסטרטגיית פעולה יציבה. ראשית, משום שהסיכוי להימנות עם 4% של הזכרים המובחרים הוא רק 4%. שנית, ככל שאחוז16 הזכרים בעדר יגדל, לא זו בלבד שהמאבק ביניהם יהיה חריף ומסוכן פי כמה, כי אם גם הפרס יהיה דל לאין ערוך. בנסיבות כאלה יהיה כדאי יותר להוליד נקבות וכך חוזר חלילה.

מאבקי מחזרים
ככל שנכון הדבר שהאסטרטגיה האבולוציונית היציבה דואגת לשיוויון מספרי בין המינים, רבות העדויות כי הטבע בכללותו נוטה לשוביניזם זכרי, וזאת על-פי כללי משחק שנקבעו כבר ברמת הביצית הנקבית והזרעון הזכרי.
כאמור, הפער בין הזכר לנקבה בא לביטוי, בדרך-כלל, בעובדה שמספר הנקבות שיכול הזכר להפרות מוגבל רק ע״י אונו וזריזותו, בעוד שהנקבה, מהרגע שהופרתה, יוצאת ממשחק הזיווגים בכל תקופת הריונה ובמקרים רבים אף בכל תקופת הגמילה. כיוון שכך יהיה זה מעשה נואל מצד הנקבה להזדווג עם כל בר-בי-רב. כמשקיע נבון היא תערוך מיון קפדני, שבסופו תעתר רק לבולטים ולחסונים מבין מחזריה, היכולים להוריש כשירות דארוויניסטית מירבית לצאצאיה, נושאי הגנים שלה. מיון קפדני זה מוסדר באמצעות מאבקי כוח וראווה בין הזכרים, אשר בסיומם זוכים המנצחים בפרס המיוחל – הזכות להפרות את הנקבה.
היה זה דארווין שחילק את המיון המיני לשני סוגים: הבירור התוך-מיני – שבו הזכרים מפגינים תוקפנות ומפעילים כוח כדי לזכות בחסדי נקבה; סוג זה מביא, באמצעות הבירור הטבעי, להתפתחות שרירים, מידות גוף, תוקפנות וכלי זין, כגון: טפרים, שניים, קרניים וכיו״ב, המגבירים את כושר הלחימה של הזכרים.
סוג שני, הבירור הבין-מיני, שם את הדגש לא על הכרעת היריב אלא על משיכת תשומת ליבה של הנקבה. סוג זה מביא באמצעות הבירור המיני להתפתחות צבעים ססגוניים, מרקמי נוצות, תצוגות של כרבולות, זנבות מפוארים וכיו״ב מרכולות שיש בהן כדי לרכוש את לב הנקבות.
דוגמא אופיינית לבירור תוך-מיני היא מאבק הזכרים ממין הקרפדה האירופית המצוייה. אף שמאבקים אלה אינם מסתיימים, בדרך כלל, בפציעה או במוות, הם מחזיקים בשיא המשך: עד 12 שעות ברציפות, שבסופן מותשים היריבים לחלוטין. למעשה מתחילה ההתמודדות כבר בדרך לבריכה, המשמשת אתר בלבדי לזיווגים. מאחר שהבריכה שורצת זכרים הממתינים לבואן של הנקבות, נוטים זכרים רבים ליירט את הנקבות העושות דרכן לבריכה: הם מזנקים עליהן, לופתים את גופן בגפיהם הקדמיות ורוכבים על גבן עד לבריכה. אלא שהמסע אינו כה נינוח כפי שהוא נשמע. לכל אורך הדרך מתנפלים על הרוכב זכרים להוטים ומנסים להפילו. הרוכב, מצידו, מפעיל את גפיו האחוריות ובועט ללא הרף על ימין ועל שמאל. אך כל זה אינו אלא הקדמה. בהגיעו סוף-סוף לבריכה מתחילה המהומה הגדולה: חמישה, ולפעמים אף עשרה זכרים, נאבקים על חסדי הקרפדה המסכנה ונערמים עליה בערימה גדולה. וכך קורה לא אחת שהזכרים הנלהבים נאבקים על גוויה של קרפדה שמתה תחתם בחנק.
לעומת חינגת האלימות של הבירור התוך-מיני, נושאים המאבקים של הבירור הבין-מיני אופי טכסי של קרבות אבירים.
דוגמא מאלפת לבירור בין-מיני, שכל תכליתו לקבוע את סדר הדומיננטיות בקרב הזכרים (ומכאן – את זכות הראשונים להפריית הנקבות), היא התופעה של ״זירת הקרב״.
״זירת הקרב״ היא אזור מוגבל בשטחו, נטול כל חשיבות מבחינת משאבי מזון, המשמש אתר התמודדות והזדווגות גם יחד. התמודדויות במסגרת ״זירת הקרב״ נהוגות בקרב מינים רבים של חרקים, דגים ויונקים. אחד המרתקים, וגם היותר נחקרים, מבין מופעיה מוצג ע״י השכווי (תרנגול בר) האמריקני.
זירת הקרב, המשתרעת על שטח של 10 דונם לערך, מאוכלסת ע״י 400 זכרים בקירוב, המתמודדים על הבעלות על מגרשי רבייה בני 10 עד 100 מטרים רבועים. אלא שלא כל המגרשים שווים, ערכם עולה ככל שהם קרובים למרכז הזירה, שכן הנקבות מעדיפות את הזכרים שמגרשיהם נמצאים ב״מקום טוב באמצע״.
התוצאה: פחות מ-10% מהזכרים, אלה שרכשו את המגרשים המרכזיים, מפרים יותר מ-75% מהנקבות הנוהרות מכל עבר לביקור חטוף בזירת הקרב. ביתרת הנקבות, אם בכלל, מתחלקים בעלי המגרשים בפריפריה.
לכאורה ממלאות הנקבות תפקיד פאסיבי, אבל למעשה כל המאבק העיקש הזה על המגרשים הזעירים הוא משחק שאת חוקיו קבעו הנקבות ע״מ לאתר להן את בני הזוג המוצלחים והחסונים ביותר, בבחינת ״ישחקו הנערים לפנינו״, בדומה לקרבות האבירים הססגוניים. בהבדל אחד, מאחר שההתמודדות נערכת במסגרת הבירור הבין-מיני, אין בה כמעט אלימות וכל כולה צבע, צורה וקול. הזכר מנפח את החזה האלסטי שלו (בעל תכולה של 5 ליטר), מציג לראווה את הנוצות הלבנות שבצווארו ומרחיב את המסרקות הצהובים שעל עיניו. לפתע, פעמיים בזו אחר זו, הוא מגביה את שק החזה שלו ושומטו: תוך כדי הרמת השק הוא מותח את כנפיו אחורה, לאורך הנוצות הנוקשות שלצידי חזהו. התוצאה: סדרה של שריקות עמומות ורעשים קטועים, הנשמעים למרחק של מאות מטרים. קולות אלה, המלווים תצוגה מרהיבה של צבעים וצורות – הם הם כלי הנשק המשמשים את המתמודדים על המגרשים המרכזיים.

לאהבה יש מחיר
כבר עמדנו על כך שהמנגנון המרכזי בטבע, המכוון ומשכלל את התכונות של בעלי החיים, הוא הבירור הטבעי. מכאן נובע שכל תכונה, החל בקרני ראמים וכלה בצבעי צפורים, אשר מסיבה כלשהי הפכו מקור משיכה לנקבות, תתפתח בקצב מהיר ותתפשט בכל האוכלוסיה. שהרי הנקבות יעדיפו להזדווג עם בעלי תכונה זאת ויעבירו אותה לצאצאיהן.
לחץ אבולוציוני זה לטובת תכונות פיסיולוגיות מסויימות הוא כה חזק, עד כי במקרים מסויימים התפתחותן פוגעת בכושר ההשרדות של בעלי התכונה המועדפת. דוגמא מובהקת להתפתחות כזאת באה לביטוי במשפחת הפאסיונים. הזכרים בולטים בשלל צבעיהם ובזנב הנוצות האדיר באורך של עד 160 ס״מ (הפאסיון המלכותי), אותו הם פורשים ומציגים לראווה לעיני הנקבה – בכדי לכבוש את ליבה. הבעיה היא שזנב אדיר זה, שכה מיטיב להמיס את ליבן של הנקבות, מסרבל את תנועתו של הפאסיון ומפחית מכושרו לחמוק מטורפים. לפנינו, איפוא, מקרה קלאסי של התנגשות בין שני היבטים של הכשירות הדארוויניסטית: הגברת כושר ההתרבות, מצד אחד, ושכלול כושר ההשרדות – מצד שני. במקרה של הפאסיון דומה כי גברה ידו של הראשון.17 מקרה דומה של סתירה בין שני עקרונות היסוד של הכשירות הדארוויניסטית, אך עצוב יותר, ארע אצל האיל האירי שנכחד לפני 11,000 שנה לערך. איל גדול מימדים זה פיתח כפות קרניים אדירות, שמוטתן הגיעה לארבעה מטרים ומשקלן ל-45 ק״ג!
הפולמוס על הסיבות שהביאו להכחדתו של האיל האירי נטוש כבר 200 שנה לערך – האם נכחד בהפשרה הגדולה של הקרחונים, או שמא להיטותו לשאת חן בעיני הנקבות, במסגרת המאבקים הבין-מיניים והתוך-מיניים, הביאה את קרניו למימדים מפלצתיים ובדיעבד קטלניים?
ממעקב אחר התנהגותם של קרובי משפחתו המודרניים של האיל האירי, שולל ג׳אי גולד את הגרסה לפיה התפתחו קרניו ככלי לחימה במאבק התוך-מיני. לדעתו הן שימשו למטרות תצוגה טקסית בין הזכרים, במסגרת המאבק הבין-מיני. גולד מצביע על כך שקרוביו העכשווים של האיל האירי מסובבים את ראשם מצד לצד כדי להציג לראווה לפני יריביהם את מלוא הדר קרניהם.
תנועה סיבובית כזאת היתה עלולה לנקוע את צוארו של האיל האירי, מחמת משקלן הרב של קרניו. כאן אולי ההסבר לעובדה שכפות הקרניים של האיל האירי היו מאורגנות בזווית כזאת, שהיו מוצגות במלוא הדרן בשעה שהראש מופנה הישר לפנים.

תחרות השאגות של האיל האדמוני
כך או כך, זנבו הארוך של הפאסיון וקרניו הגדולות של האיל האירי (אם אכן הן אחראיות להכחדתו) מעלות שאלה יסודית יותר הנוגעת למאבק הבין-מיני: מה יש בקרניים הגדולות, בזנב האדיר, בנוצות הססגוניות וכיו״ב, הכובשים את ליבן של הנקבות? מה היתרון הכשירותי הטמון בקישוטים אלה, המביא את הנקבות להעדיפם על-פני זכרים פחות גנדרניים? הדעת הרווחת היא שבשלב הראשון של התפתחותם, נשאו קישוטים אלה יתרון השרדותי כלשהו, אשר תרם לכשירות הדארוויניסטית של הצאצאים. למשל, יתכן שפאסיונים בעלי זנבות קצת יותר ארוכים, היו זריזים ומיומנים יותר בתמרוניהם מאשר בעלי הזנבות הקצרים. וכך, מרגע שזנב ארוך יותר הוכר כיתרון, זכו בעליהם להעדפה אצל הנקבות ולפיכך אחוז צאצאיהם באוכלוסיה גדל בהתמדה. עכשיו, ככל שהמאבק על הגישה לנקבות נעשה עז יותר, הפך הלחץ להארכת הזנב למטרה בפני עצמה, למרות שהיתרון ההשרדותי שהוא ייצג בראשית התהליך – נעלם מזמן.
לחוקר הישראלי, פרופ’ אמוץ זהבי, הסבר משלו להתפתחותן של התנהגויות ותכונות המסכנות את בעליהן או פוגעות בכושר השרדותם. לדעתו, הזנב המפואר לא נועד להכות בסנוורים את הנקבה, כי אם להבליט לעיניה את תועפות חוסנו בדרך השלילה: למרות המגבלה הקשה שבזנב ארוך ומסורבל זה, אני חי וקיים.
מסקנה – אני חסון ומוצלח מבעלי הזנבות הקצרים יותר. הוא הדין באותם בני בקר הרוקעים בפרסותיהם בקרוב אליהם טורף. אליבא דזהבי, אין בכך מעשה אלטרואיסטי שנועד להזהיר העדר מפני הסכנה, אלא מסר גלוי ושחצני כלפי הטורף: אם חשקה נפשך בטרף, בחר לך קרבן חלש ופחדן ממני.
השאלה אם מסר מינכהוזני זה (כאותו רץ מופלא בספר עלילותיו של הברון מינכהוזן, שהיה נאלץ לקשור משקולת כבדה לרגליו בכדי להאיט את מאוצו המהיר) נתפש ע״י החיות בטבע, נותרה לפי שעה פתוחה. למעשה, גם ההסברים האחרים על תפקיד הצורות והצבעים הססגוניים במשחק הבין-מיני הם בגדר הנחות שלא הכל שלמים עימן. מה שברור הוא שיש בקישוטים אלה יתרון, שאם לא כן לא היו מתפשטים ומתבססים באוכלוסיה, ובודאי שהיו נכחדים מחמת החסרונות ההשרדותיים לכאורה שהם נושאים.
אחד היתרונות הבולטים שיש לחלק מקישוטים אלה, הוא ביכולתם להכריע את ההתמודדות בין הזכרים היריבים בכל הקשור להגמוניה ולמקומם בהיררכיה ובכך גם להקנות להם גישה מועדפת לנקבות ולמקורות מזון – בלי להזדקק לעימות פיסי, שסכנתו מרובה למתמודדים.
דוגמא מאלפת לשימוש ב״קישוט״ קולי היא שאגתו של האיל האדמוני. האיל האדמוני, החי באיזורים שונים באירופה, הנו חיה מרשימה, בעלת מוטת קרניים גדולה (120 ס״מ), המתפצלות לסעיפים רבים וחדים. בכל ימות השנה חיים הזכרים בשלום אלה עם אלה, אך בחודש אוקטובר, בהפתח עונת הייחום, מתלהטות הרוחות והם הופכים יצורים אלימים ביותר. למעשה ניטש המאבק על השליטה בחלקות מרעה משובחות, שיש בהן כדי למשוך אליהן מספר גדול של נקבות. החשבון פשוט ואכזרי: מי שאינו מצליח לתפוס חזקה על שטח מרעה – אין לו זכות גישה לנקבות, ועליו לחכות לעונה הבאה כדי לנסות שוב את מזלו.
בשעת המאבק משלבים הזכרים את קרניהם אלה באלה ומנסים להסיג האחד את רעהו לאחור. גורל הקרב תלוי בעוצמת שרירי הצואר והירכיים, אך לא פחות מזה גם בעבודת רגלים זריזה ומיומנת, העושה שימוש נבון בפני הקרקע והמביאה את היריב לאבדן שיווי משקלו.

הפרס, כמובן, הוא הזכות להרביע כ-20 נקבות. בפועל, התוצאות קלושות יותר. אחדות מהן אינן בוגרות עדיין ואחדות לא מצליחות להתעבר, כך שמספר צאצאיו (בהתחשב בתמותה הטבעית) לעונה עשוי להגיע רק לחמישה. כדי לזכות בפרס זה משקיע הזכר אנרגיה אדירה ומהמר הימור מסוכן על בריאותו. למעשה מבלים בעלי החלקות את מרבית זמנם בסימון גבולות הטריטוריה שלהם ובמאבק בלתי פוסק בפולשים המנסים להשתלט על נקבותיהם. בדרך כלל מקדישים האילים 6 שעות ביום למרעה. אך בעונת הייחום, הנמשכת כחודש ימים, הם יכולים אך בקושי להרשות לעצמם הפסקת אוכל בת חצי שעה. מאחר שכך ניזונים הזכרים בעיקר מעתודות השומן שברקמותיהם, והם מאבדים במרוצת חודש אוקטובר הרבה ממשקל גופם. למעשה, עליהם לכלכל את מאבקיהם בשום שכל, שכן הם עלולים להתיש כוחם כליל עוד לפני שמגיעות הנקבות לייחומן.
המאבק עצמו, אף שהוא מתבסס בעיקר על דחיפות הדדיות בקרניים שלובות, מסתיים לעתים קרובות בשברים, בפציעות קשות ואף בריסוק קרניים (שפירושו – אבדן כושר ההתמודדות). כאן מגיע חלקם של ה״קישוטים״. במקום להסלים כל עימות למאבק פיסי, התפתחה במרוצת האבולוציה שיטה מונעת, המכריעה בסוגיית המנצחים והמפסידים על-פי תדירות השאגות. למשל, כאשר זכר חופשי חושק בהרמון של יריבו, הוא פותח בשאגות לעברו. אם, לדוגמא, שאג ארבע שאגות בדקה ובעל ההרמון השיב לו בשבע שאגות, מכיר המתמודד בנחיתותו ויוצא לחפש את מזלו בשדות אחרים. אם קצב שאגותיהם שווה פחות או יותר מתחילים השניים לצעוד זה במקביל לזה, תוך שהם שואגים במלוא ריאותיהם. ואם גם בשלב זה אף אחד מהם לא מוותר – מתפתח הקרב הבלתי נמנע.18 כאן, כמובן, יכולה להשאל שאלת-תם: אם ניתן להכריע קרב על-ידי שאגות, מדוע לא עודד הבירור הטבעי הופעתם של אילים בעלי כישורים קוליים מופלגים, על חשבון שרירים וקרניים ראוותניים, הדורשים השקעת אנרגיה מרובה? התשובה: משום שההונאה לא יכולה להוות אסטרטגיה אבלוציונית יציבה. שכן, במוקדם או במאוחר תוכיח המציאות שלשקר אין רגליים. מכל מקום, המחקרים הראו שכושר השאגה של האילים האדמוניים תואם בקירוב גבוה את כושר לחימתם.
אף על פי כן – הונאה
עם זאת, אף שהתנהגות נוכלית אינה יכולה להוות אסטרטגיית פעולה יציבה לאוכלוסיה בכללותה, לא נעדרים בטבע מעשי הונאה. אין זאת שהטבע מגלה סובלנות כלפי גומחות ״נוכליות״ כאלה, בתנאי שאינן מתפשטות יתר על המידה, עד כדי שיבוש האיזון בקרב האוכלוסיה בכללותה. בקרב קרפדות העצים, החיות באמריקה הצפונית, מכוונים הזכרים את הנקבות לעברם ע״י קריאות קצובות. אבל, אחד לשבעה זכרים לערך, אינו קורא, אלא תופש עמדה ליד זכר רעשן כזה וממתין בדממה גמורה. בהתקרב הנקבה הוא קופץ על גבה ומסיים עניינו עימה ללא שהיות, בלי להשחית זמנו בקריאות מייגעות.
דוגמא מתוחכמת יותר של הונאה נהוגה ע״י הסלמנדה הנמרית האמריקנית. תהליך ההפרייה מתנהל בדרך זאת: הזכר צועד בראש ומכוון בטלטולי זנב קלים את הנקבה הצועדת בעקבותיו. כאשר היא מוכנה להפרייה היא נוגעת קלות בראשה בזנבו. מגע זה מהווה לזכר אות לשחרר מפתחו מיכל זרע על הקרקע. הנקבה עוברת מעל המיכל ומכניסה אותו לקירבה דרך הפתח הנקבי שלה. ספור פשוט זה לא היה שווה אזכור, אלמלא התערבותם של מספר נוכלים, המנצלים את הסיטואציה האינטימית לצרכיהם הם. נוכל כזה מיירט את הזוג ונכנס בין הזכר לנקבה, לאחר זמן הוא נוגע קלות בזנב הזכר שלפניו, והלה, כמובן, מטיל את מיכל הזרע שלו על הקרקע. הנוכל הצועד בעקבותיו, מניח על מיכל קודמו את מיכלו שלו. הנקבה שצועדת בעקבותיהם נוטלת לה את המיכל העליון ומותירה במקום את מיכלו של מחזרה המקורי.
סוג אחר של הונאה רווח בקרב חרקים מסדרת ארוכי הכנף (Black Tipped Hanging Fly), החיים בצפון אמריקה. תחת להתעסק בחיזורים מייגעים ובמאבקים מתישים, קונה חרק זה את חסדי המיועדת לו תמורת אתנן. הזכר מביא לכלתו כנימה דשנה, ובעוד היא עסוקה בבדיקת המתנה העסיסית, הוא מזדווג עימה. אך לא תמיד צולחים מעשיו. אם השי קטן ודל מדי, דוחה אותו הכלה הבררנית מעל פניה ומסרבת להזדווג עמו. אולם גם הוא לא טומן ידו בצלחת. ההזדווגות אורכת בממוצע 23 דקות, אך אם הזכר זריז דיו והוא מסיים מעשיו לפני שהיא גומרת לסעוד, אין הוא מהסס לחטוף את הציד מפיה ולהשתמש בו כמתנת כלולות חדשה לנקבה אחרת. אכן, ההגינות אינה שורה בקרב חרקים אלה, מה גם שזכרים רבים מנסים לרכוש אלה מאלה שי כלולות בחטיפה. לשיטה זאת, כמובן, יתרון ברור. כל גיחת ציד עלולה להסתיים בגפיהם השעירות של עכבישים, אשר רשתותיהם פרושות בכל מקום. מכאן, הרבה יותר בטוח לחטוף טרף מוכן מאשר לשחר אחריו. החטיפה נעשית בשתי שיטות: בעימות אלים עם זכר אחר, או ע״י התחזות לנקבה. המתחזה נענה לריח הפרומון (הורמון מין) שמשחרר הזכר מבלוטותיו (כדי למשוך לעברו את הנקבות), מאותת לעברו בכנפיו – כאות לכך שהוא מתרצה לתשורתו ומוכן, כביכול, להזדווג עימו – ובעוד הראשון מנסה נואשות לממש את האתנן, חוטף המתחזה את הטרף ונעלם.
מבין מגישי התשורות אין ספק שפרס ״הנקמה המתוקה״ מגיע למין של זבובונים ממשפחת האמפידה. כמקובל בקרב מינים רבים ממשפחה זאת הנקבה נוהגת לטרוף את בן זוגה. כדי לעקוף סכנה זאת נוהג הזכר להביא עימו שי ולנצל את התעסקותה במזון כדי לממש את צו הטבע בבטחון יחסי. (אגב, יש מינים הקונים לעצמם בטחון נוסף באורזם את השי בחוטי משי, דבר המאריך את משך התעסקותה של הנקבה במתנת הכלולות). אלא שהשי אינו אלא אריזה חלולה. לאחר שהזכר לוכד טרף הוא מוצץ את לשדו ואח״כ עוטף את הקליפה היבשה באריזה עבה של חוטי משי. הנקבה מקבלת מידו את השי בהתלהבות וניגשת לאלתר למלאכת הפרימה המסובכת. אך עד שהיא מסיימת, עושה הזכר את שלו ומותיר את הנקבה הטורפנית עם גוויה צמוקה של חרק.19
הקוקיה הרצחנית
אם לסכם דוגמאות אלה, דומה כי הקו המשותף להן הוא יסוד הטפילות שבהן. משמע, תחת לעמול ולהשקיע אנרגיה מרובה להשגת מזון ונקבות, או להקדיש זמן לגידול צאצאים, נהנים הטפילים מן ההפקר. למשל, בקרב מינים מסויימים של נמלים, המלכה אינה שבה לקינה בתום מסע הכלולות, שם נכון לה מאבק באחיותיה המלכות. תחת זאת היא נוחתת ליד מושבת נמלים ממין אחר, מתחזה כמתה ומניחה לנמלים הפתיות לגרור אותה לתוך קינן. משחדרה פנימה, היא הורגת את המלכה המקורית ומניחה לפועלות המקומיות לגדל את צאצאיה הנושאים גנים זרים להן.
הדוגמא היותר מוכרת של טפילות היא, כמובן, זאת של הקוקיה, המטילה את ביציה בקן של צפור פונדקאית, ומעבירה אליה את עול גידולם.
כמספר מיני הקוקיות, כן מספר שיטות ההונאה שהן נוקטות כדי להוליך שולל את הפונדקאית. למשל, זכר הקוקיה חג במתכוון סביב קן הפונדקאית ובעוד היא יוצאת לקראתו כדי לעמוד על כוונותיו, מתגנבת נקבתו לקן ומטילה בו בחופזה את ביציה. אך בכך לא מסתיימת תכנית הפעולה השטנית של הקוקיה. אך בוקע גוזלה מביצתו הוא מפעיל שיטה מדהימה לחיסול הביצים ו/או הגוזלים של הצפור המארחת, כדי להבטיח לעצמו את מלוא המזון של ההורים: כל ביצה או גוזל הנלחצים אל גופו הוא עומסם על גבו ומטפס עימם עד שפת הקן. בהגיעו לשם הוא שומטם החוצה אל מותם הוודאי. אצל מין אחר של קוקיה הגוזל משסע במקורו המעונקל את גוזליה של הצפור המארחת עד שהוא נותר בודד בקן.
כאמור, הטפילות אינה יכולה להוות אסטרטגיית פעולה יציבה לכלל הפרטים באוכלוסיה, שהרי אם כולם טפילים אין על-חשבון מי לחיות. ובכל זאת, לנוכח היתרונות העצומים שיש לטפילות, מוזר שהיא לא פשטה והתבססה – ולו חלקית – בקרב כל המינים. ולהיפך – מאחר שהטפילות מייצגת מבחינתו של המארח בזבוז מוחלט של השקעתו, שאינה תורמת דבר וחצי דבר להפצת הגנים שלו, תמוה הדבר איך לא עודד הבירור הטבעי התפתחותם של מנגנונים אנטי-טפיליים (דוגמת הנוגדנים של מערכת החיסון), המסוגלים לשים לאל את תכניתם השטנית של הטפילים. אך על זאת מאוחר יותר.
חשוב לציין שיש מקרים בהם יוצרים הטפילים קשר סימביוזי עם מארחם, שבו שני הצדדים נהנים. למשל, אצל אחד מחמשת המינים של צפור הבקר האמריקנית, מתקיימת סימביוזה מעניינת ביותר בינם לבין מארחיהם. מסתבר שהצפורים המארחות סובלות קשה מעונשו של חרק המתחפר בבשרם של הגוזלים ומביא למותם. יש הבונות את קיניהן בקרבת מושבות של צרעות וזוכות מהן להגנה מפני החרק. שיטת הגנה אחרת היא לאפשר לצפור הבקר להטיל ביציה בקיניהן. הגוזלים האורחים, בעלי החיוניות הגבוהה, מנקרים מבשר מארחיהם את ביצי החרקים ואף תוקפים בחמת-זעם את החרקים הבוגרים המנסים להטיל ביציהם בקן. בדרך זאת קונה לעצמה המארחת הגנה מירבית מפני החרקים, תמורת מעון ומזונות לגוזלים הזרים. למעשה אפשר לראות בסימביוזה כזאת מעין גרסה של ״אלטרואיזם הדדי״ (מונח שטבע טריברס), השכיח למדי בטבע בין פרטים שאין ביניהם קירבת משפחה. על-פי ה״אלטרואיזם ההדדי״, או בנוסחו העממי: ״גרד לי ואגרד לך, צד א׳ גומל טובה לצד ב’ ומבטיח בכך החזר טובה מצד ב’. למשל, בקרב הבאבונים קורה לא אחת שיריבים המתמודדים ביניהם מבקשים את עזרתו של באבון שלישי, ומשיבים לו מידה כנגד מידה כאשר הוא נקלע לצרות.

הצבע הוא המסר
אך הבה נחזור לסימן השאלה שהצבנו סביב תצוגות הראווה ומפגני הצבע הססגוניים שמפעילים הזכרים במסגרת הבירור הבין-מיני. יש הרואים בהם מנגנונים מתוחכמים לקביעת מקומם של הזכרים בסולם ההיררכיה, תוך עקיפת עימות פיסי אלים שסכנתו גדולה והצורך אנרגיה מרובה. בחלקם, בעיקר אותם מפגנים ותצוגות ראווה המקדימים קשר מונוגמי בין הזכר לנקבה, תפקידם לאלץ את הזכר להשקיע זמן ואנרגיה להשגת הנקבה, כך שהסתלקותו בתום ההזדווגות לא תהיה כדאית לו. החוקרים רובין באקר וגאוף פרקר גורסים שהצבעים הססגוניים מיועדים להעביר מסר לטורפים ולא לנקבות. הצבעים העזים, טוענים שני חוקרים אלה, באים להעיד על בעליהם שהם ניידים ביותר או בעלי ראייה טובה מאור, ולפיכך יהיה זה בזבוז זמן מצד הטורף לרדוף אחריהם. אדרבא, אומרים צבעים אלה, חפשו את סעודתכם בקרב הנקבות נטולות הצבע, שעפי״ר הן גם קטנות וחלשות יותר. באקר ופרקר לא מסבירים מדוע צריכות הנקבות להשלים עם אגואיזם שוביניסטי זה מצד הזכרים, ומדוע לא דאג הבירור הטבעי לעודד צבעוניות גם בקרב הנקבות. והלא כבר נוכחנו כי הנקבות מיטיבות לדאוג לאינטרסים שלהן, עד כדי שלילת זכות ההפרייה ממרבית הזכרים, וזאת בכדי להבטיח לעצמן את הגנים המובחרים של החסונים והמעולים שבהם. כך, למשל, נוהגת נקבת פיל הים לזעוק בקולי-קולות מאימת שזכר להוט מתקרב אליה. זעקותיה ההיסטריות מחוללות מהומה רבתי בעדר ומביאות את הזכרים להתלהטות יצרים. הם נאבקים אלה באלה בשצף קצף בעוד הנקבה ממתינה בצד למנצח כדי להזדוווג עימו. ואכן, היה זה החוקר הדגול ר.א. פישר, אשר ב-1930, בספרו על התאוריה הגנטית של הבירור הטבעי, העניק בסיס מתימטי להשקעה היחסית של בני הזוג בצאצאיהם. והכלל שגזר היה, כי הנקבות בררניות יותר בבחירת בן זוגן מאשר הזכרים. שכן השקעתן בהקמת צאצאים גדולה יותר, לפיכך הן עלולות להפסיד יותר אם לא יבחרו בבעל הגנים החסון והמוצלח יותר.

סוף מעשה במחשבה תחילה
אנו רואים איפוא שהטבע משופע מכאניזמים ממכאניזמים שונים, פיסיולוגים, מורפולוגים, התנהגותיים וכו’, שתכליתם להקנות לפרט כושר השרדות גבוה יותר ויכולת הסתגלות טובה יותר לסביבה, למען יוכל לממש את מטרת קיומו: הקמת צאצאים כאמצעי להרחבת מאגר הגנים שלו ולהפצתם ברבים. ומאחר שהאוכלוסיה גדלה בקצב הרבה יותר גדול מאשר משאבי המזון, כפי שהראה בשעתו מלטוס, ניצב מעליה מנגנון הבירור הטבעי, כחרב מתהפכת הגוזרת כליה על כל החלשים והפחות מוצלחים ויעילים, אשר אינם עומדים במאבק הקיום האכזרי בין טורפים לנטרפים, המתנהל 24 שעות ביממה. במילים אחרות, הבירור הטבעי יותיר בחיים רק אותם פרטים שפיתחו ושיכללו תכונות ומכאניזמים מוצלחים ויעילים יותר מזולתם. אך יעיל יותר אין פירושו דווקא אלים או זללן יותר, או אפילו בעל כושר שיכפול עצמי גבוה יותר. למשל, ב-1950 הובא לאוסטרליה נגיף המייקסומה במטרה למתן את ההתרבות שלוחת הרסן של אוכלוסיית הארנבות. אלא שהנגיף היה כה אלים עד כי חיסל את הארנבות (שנדבקו ממנו) עוד טרם היה סיפק בידו להשתכפל בכמות מספקת ולהדביק ארנבות אחרות. במילים אחרות, במוות המהיר שהביא על פונדקאיו – הביא גם כליה על עצמו. ואמנם, זן מתון יותר של נגיפים, שהתפתח במרוצת הזמן, ירש את קודמיו האלימים והביא לחסולה המלא כמעט של אוכלוסיית הארנבות. מסקנה: הנגיף המתון יצג במקרה זה אסטרטגיית פעולה שונה יותר, אשר איפשרה לו לנצל ביעילות את משאביו ומנגנוני השכפול של הפונדקאי שלו. או ניקח דוגמא אחרת מקרב הטורפים. טורף טריטוריאלי שיטרוף טרף מכל הבא ליד, בלי לעשות את חשבון העתיד, יפגע בסיכויי ההשרדות של צאצאיו, ולפיכך תפוצתו של גן “פזיז” זה תצטמצם ותלך, מכאן, הבירור הטבעי צריך לעודד במצב כזה התפתחותה של התנהגות גנטית שקולה ומחושבת יותר. ואמנם, מעקב אחר להקות זאבים הראה כי הם מעדיפים טרף צעיר מאוד או מבוגר מאוד, המייצגים פרטים בעלי פוריות נמוכה.
האם יש לראות בכך העדפה משקית נבונה או נטייה פשוטה וטבעית לבחור בטרף קל יותר לציד? שוני המעקב לא נותנים תשובה חד-משמעית. מכל מקום, ברור שבעלי חיים מביאים בחשבון את משאבי הסביבה, בפועל וגם בכוח, בתכנון פעילותם. ידוע, למשל, שמספר הביצים שמטילות הצפורים שונה בעונה שחונה מאשר בעונה גשומה. יש עדויות רבות לכך שמספר הביצים בקן תואם את מספר הצאצאים המירבי שההורים יכולים לספק להם מזון. וכבר הוכח כי צפורים המקפידות על הטלת כמות מיטבית של ביצים, מצליחות לגדל יותר צאצאים מאשר צפורים המטילות יותר (או פחות) מהמספר המיטבי.
אולם הערכה נכונה של משאבי הסביבה מהווה רק שיקול אחד בעבור הצפור. השקול האחר הוא שיעור הסכנה האורבת לצאצאיה מטורפים. משמע, ככל שהטורפים מרובים יותר וסכנתם גדולה יותר, עליה להטיל יותר ביצים. שכן מספר גדול יותר של ביצים20 מגביר את סיכויי הפליטה להשרד. מצד שני, בכל יצירת ביצה משקיעה הצפור משאבים לא מבוטלים. מכאן, המספר המיטבי של הביצים צריך לענות על מספר מדדים: מספיק ביצים כדי להבטיח השרדות הצאצאים, אבל לא יותר ביצים מכפי יכולת הסביבה וההורים לקיים.

מפקד אוכלוסין
מקובל עלינו שאוכלוסיה צפופה בשטח נתון, שהוא, מדרך הטבע, בעל משאבים מוגבלים, פוגעת בכשירותו הדארוויניסטית של כל פרט באוכלוסיה, באשר המחסור במזון יעלה את אחוז התמותה, דבר שיביא לדילול הצפיפות באוכלוסיה. שיווי משקל זה בין גודל האוכלוסיה לבין משאבי הסביבה, נשמר גם באמצעות מנגנונים אחרים. למשל, ככל שהאוכלוסיה גדלה, מתרבים גם מספר הטורפים, שכן היא מהווה מקור מזון עשיר ונוח להשגה. אבל ככל שהטורפים מתרבים, השגת המזון נעשית קשה יותר, ומספרם מצטמצם. בדרך זו דואג הטבע לאיזון טבעי בין טורפים לנטרפים. אך נראה שהטבע לא הסתפק באיזונים טבעיים אלה, וכי לצידם פועלים מכאניזמים נוספים לפיקוח על הילודה, הזוכים, ככל הנראה, לעידוד מצד הבירור הטבעי. אין זאת שהתדלדלות מקורות המזון ועליה בצפיפות האוכלוסיה מפעילות מנגנונים הורמונליים, המגבירים את התוקפנות ההדדית בין הזכרים וגורמים לירידה דרסטית של הפריון בקרב הנקבות. למשל, בגבור הצפיפות באוכלוסיית מין נפוץ של עכברים בנורבגיה, מתפתחות התנהגויות חריגות של פתולוגיה חברתית, הנראות כהתאבדות המונית. במקביל לכך לא משלימות הנקבות בניית מרבצי ההמלטה שלהן ותמותת ולדותיהן מגיעה עד 90%. בקרב יונים גורמת הצפיפות לזכרים להפריע זה לזה מלהזדווג. תופעה של אכילת צאצאים רווחת במינים רבים של בעלי חיים.
ככלות הכל, משפוחת המזון הופכים הוולדות למנת הברזל הקרובה והזמינה ביותר. בקרב הטרמיטים הקניבליזם הוא צורך חיים. כל אימת שנוצרת אוכלוסיה גדולה מדי של צאצאים – הם נאכלים ע״י הפועלות. הללו טורפות גם את הזכרים ומונעות בכך יציאתם למפגש ההפריה עם המלכה. יש ותופעות חריגות אלה משבשות באורח חמור, ולעתים גורלי, את הסדר והאיזון בקרב אוכלוסיות המינים. שיבושים אלה אפשר היה למנוע לו ערכו בעלי החיים מפקד אוכלוסין תקופתי בכדי לפקח על הילודה. והנה, עד כמה שהדבר נשמע מפתיע, במיוחד לנוכח כשלון מאמצי הפיקוח על הילודה בקרב בני האדם, יש הטוענים לקיומם של מנגנונים כאלה בקרב בעלי חיים.
על-פי התיזה של וין-אדווארדס, תצוגות הראווה של הזכרים באות בראש וראשונה להעריך את מספר הזכרים הזמינים, ולהכריע בהתאם לכמה מהם תינתן זכות ההפרייה בכל עונה ועונה3 לדעת וין-אדוארדס הפריית הביצית היא אך מטרה משנית של המאבקים הבין-מיניים והתוך-מיניים. מטרתם המרכזית לוודא שהאוכלוסיה לא תעבור את הצפיפות המיטבית ביחס למשאבי המזון הנתונים.
לתצוגות אלה (Epideictic Display), הממלאות תפקיד של מפקד אוכלוסין, צורות ביטוי שונות ומגוונות, לדידו של וין-אדווארדס, מטרת ההתגודדויות ההמוניות של זרזירים לקראת ערב, או של ענני היבחושים המחוללים אלה לעומת אלה, היא לקיים מפקד אוכלוסין, בשיטה פשוטה ויעילה: צפיפות המתכנסים מעבירה מסר בלימה (או שחרור) למנגנוני ההפרייה של הפרטים.
גרסתו של וין-אדווארדס עוררה התנגדות לא מעטה בקרב החוקרים, בעיקר משום שהיא מאלצת בעקיפין לאמץ תיזה מיושנת ודחויה, לפיה פועל הבירור הטבעי לטובת המין בכללותו ולא רק לטובת הפרט. שכן, אם את ויתורה האלטרואיסטי של הדבורה על פוריותה ניתן להסביר בשעור הקירבה הגבוה בינה לבין אחיותיה, הרי אין להניח שעור גבוה של שאירות באוכלוסיות המוניות כאלה של יבחושים וזרזירים. מכאן שהפרטים מוותרים על הפצת הגנים שלהם לטובת המין כולו. אך הלוא זוהי בדיוק הגרסה שהאבולוציוניסטים המודרניים דוחים בכל תוקף, שכן, אם טובת המין יכולה להכתיב התנהגות לפרט, מדוע לא טובת הסוג, המשפחה, הסדרה, המחלקה וכו? וחשוב מזה: אם שם המשחק הוא ויתור על האינטרס האנוכי לטובת הכלל, יש בו הזמנה גלויה להתרבותם של נוכלים שינצלו את הפרישות המינית של חבריהם כדי להזדווג עם כל נקבה שתיקרה בדרכם. זאת ועוד, פרט המוותר על הפרייה מפחית את כשירותו הדארוויניסטית, מספר צאצאיו באוכלוסיה יקטן וילך עד שתכונה זאת של ויתור לטובת הכלל תיעלם כליל. כדי לשמר תכונה כזאת, הדואגת להפצת גנים זרים על-חשבון גנים עצמיים, יש צורך במנגנון-על, כמו התודעה או האורתוגנזה (התפישה לפיה מתנהלת האבולוציה בקו ישר ועל-פי תכנית פנימית החותרת ליעד קבוע מראש).
אלא שבקבלת מנגנונים כאלה אט נוטשים את העקרון החומרי והסיבתי של האבולוציה וגולשים לתחום המטאפיסי החלקלק שקדם לדארוויניזם.
הגן האנונימי או האורגניזם הכולי
הזכרנו כבר, בדברנו על העקרון של ״ברור שאירים״, כי העמדת תכונות ומנגנונים במבחן תרומן לכשירות הדארוויניסטית הכוללת של הפרט (כלומר, לכשירותם של שאיריו הנושאים חלק זה או אחר של הגנים שלו), העתיקה את רגש הפעולה של הבירור הטבעי מזירת הפרט אל זירת הגנים. אמרנו שאין זה הבדל סמנטי, למרות שכל ארוע המביא למות הפרט או פוגע בכושר פריונו מצמצם בדיעבד את כמות הגנים שביכולתו להפיץ. שכן, אם יחידת היסוד של הבירור הטבעי היא לא הפרט – אלא הגן, לא צריכה להיות מניעה, ברוח אמרתו המפורסמת של הולדן, להקריב חיינו להצלת חייהם של 8 דודנים שלנו (כל דודן נושא 1/8 מהגנים שלנו).
לפיכך, הוספנו, השאלה אם פרט מסויים מיעט או הגדיל את כשירותו, אינה נמדדת בהכרח ע״י מותו או ע״י מספר הצאצאים שהותיר אחריו, כי אם ע״י תרומת מעשיו להגדלת מספר הגנים שלו באוכלוסיה בכללותה.
במעבר זה של פעולת הבירור הטבעי מזירת הפרט אל זירת הגנים, טמונות השלכות מוסריות ופילוסופיות כבדות משקל, שעוד נעמוד עליהן. אך תחילה יש להבין מעט יותר זירה חדשה זאת. אנו יכולים להבין כיצד יכול דחף קיום חזק יותר או תוקפנות מוגברת לשפר את סיכוי ההשרדות של הפרט במאבקו בסביבה, באויביו וביריביו. אנו יכולים גם להבין כיצד יתפשטו הגנים האחראים לתכונות אלה בקרב האוכלוסיה, שכן ככל שהם משפרים את סיכוי השרדותו של הפרט ומקנים לו יתרון לעומת יריביו ירבו צאצאיו. אנחנו מבינים תהליכים אלה באופן חווייתי, אולם משום שהכרתנו מייחסת את דחף הקיום לאורגניזם השלם, הבועט, נושך, טורף וחומק כדי להגן על הווייתו ולשמר את עצמיותו. כלומר, הוא לא נלחם על הגנת כלייתו, גופו או זנבו, אלא על כוליותו. ברוח התייחסות זאת אנו תופשים בצורה שונה את הדחף המיני.
אנחנו נוטים לראות בו מוצר מובהק של הבירור הטבעי, שכן רק מי שפיתחו דחף כזה הותירו אחריהם צאצאים. עם זאת, אין לדחף המיני בעינינו אותו מעמד כמו ליצר הרעב ולבטח לא כמו לדחף הקיום. לראייה, זכרים רבים מנהלים אורח חיים נורמלי לחלוטין, אף אם לא הגיעו כל ימי חייהם למימוש אקט ההפרייה. יתירה מזאת, פרטים מבוגרים שאיבדו את כושר פריונם, ממשיכים להאבק על חייהם בשצף-קצף, למרות שכשירותם הדארוויניסטית (פוטנציאל ההולדה שלהם) היא אפס. מכאן התמיהה: כיצד נוכל ליחס ייחוסים כוליים אלה לגנים אנונימיים שאינם אלא סוג אחד מני עשרות אלפי מולקולות בגופו של האורגניזם? כיצד קורה הדבר שהדבורה מקריבה את עצמיותה ומשעבדת את כוליותה לטובת מולקולות אלה, המתקיימות באחיותיה, ללא כל זיקה ומגע בהוויתה שלה? וגם אם נאמץ את קו המחשבה, לפיו פועל הבירור הטבעי21 בשרות האורגניזם המורחב להפצת הכשירות הדארוויניסטית הכוללת, כיצד קנתה תכונה זו לעצמה מעמד בכורה על-פני דחף הקיום?
כדי להשיב על כך יש צורך במהפך מושגי לא קטן. למעשה אנו נדרשים לשנות את זווית הראייה שלנו.
הגנים חיים לנצח
אכן, הגנים אינם אלא מולקולות של דנ״א, אך ייחודן לעומת המולקולות האחרות בגוף, שהן מסוגלות לשכפל את עצמן. מאחר שכך, מרגע שנוצרה באקראי מולקולה כזאת לפני 4 מיליארד שנה, ב״מרק״ הקדום של מולקולות אורגניות, רק היא נהנתה ממוטציות ששיפרו את חוסנה או הגבירו את יעילותה האנרגטית. אמנם, מוטציות דומות חלו גם במולקולות אורגניות אחרות, אך שיפורים אלה אבדו עם התפרקות המולקולות (בהשפעת הסביבה העויינת), בעוד שמולקולות הדנ״א, הודות לכושר השתכפלותן, העבירו שיפורים אלה למולקולות הבת שלהן. יכולת זאת של שכפול עצמי איפשרה להן לצבור את השינויים המועילים עם כל דור חדש של מולקולות-בת, והבטיחה קצב השתכללות עקבי ומואץ תחת פעולתו הנחרצת של הבירור הטבעי.
משמע: בהתמודדות על משאבי האנרגיה המוגבלים של סביבתן הקרובה מצליחות בעלות המוטציות החיוביות לשכפל עצמן ביעילות ובמהירות רבה יותר מאשר המולקולות הפחות מוצלחות.
על-פי תסריט זה, ששחקניו הראשיים הם המקריות והאקראיות של המוטציה, מצד אחד, והכיווניות הנחרצת (מבחינת היעילות והתאמה לסביבה) של הבירור הטבעי, מצד שני, אפשר לראות בהיווצרות המנגנונים השונים של התא (הדופן, הציטופלסמה, האברונים, ריכוז הדנ״א בגרעין וכו׳) תוצאה של הצטברות שורה ארוכה של מוטציות, שכל אחת מהן תרמה לחוסנן וליעילותן של מולקולות הדנ״א. אם נמשיך בקו מחשבה זה, גם הווצרות האורגניזם הרב-תאי, על איבריו השונים – הלוע, השוטונים, הריסים, מערכת העיכול וכו’, אינם אלא שיפורים שאומצו ע״י הבירור הטבעי כמכשירים התורמים להשרדותן ולכושר שכפולן של מולקולות הדנ״א (הגנים). ומכאן אך צעד אחד לאמירה כי ״תכלית העל״ של האורגניזמים השונים, החל בנגיפים וכלה ביונקים, אינה השרדותם כפרטים, אלא שכפול הגנים שלהם. או בלשונו הפרובוקטיבית של ריצ׳רד דאווקין (בספרו, The Selfish Gene), הקוף אינו אלא מכונה המגינה על הגנים ע״י טיפוס על עצים, ואילו הדג עושה זאת ע״י שחיה. ובעצם, כל צורות החיים עלי אדמות, לרבות בני האדם, פותחו ע״י הגנים כמכונות השרדות, כלי רכב סומים המתוכנתים לשמר את המולקולות האנוכיות4 הקרויות גנים. בנוסח פחות פרובוקטיבי אפשר לומר, על משקל אמרתו המפורסמת של סמואל בטלר (״התרנגולת היא האמצעי של הביצה לייצר עוד ביצה״), כי האורגניזם הוא האמצעי של הגנים להפיץ את עצמם. למרות הלשון הלהטוטנית של משפט זה, אין הוא משולל בסיס לוגי. יש לזכור כי ברבייה מינית האורגניזם מאבד בכל דור של צאצאים 50% מזהותו הספציפית כפרט: דור הבנים נושא 50% מהגנים שלו; דור הנכדים – 25%, דור הנינים – 12.5% וכן הלאה. לעומת זאת, הגנים – בהוותם יחידת היסוד של החומר הגנטי – אינם נפגעים או משתנים משיחלוף קטעי הכרומוסומים בין תא הזרע לביצית, שכן הם עוברים מכרומוסום אחד למשנהו בשלמותם. וכך מדלגים העתקיהם מדור לדור, בהשאירם אחריהם את גוויות בעליהם ככלי רכב נטושים בצידי הכביש של האבולוציה.
להלכה, יתכן שאנו נושאים בגופנו העתקים של גנים שנוצרו לפני 3 מיליארד שנה, אשר התגלגלו בגופם של דגים, זוחלים, עופות ויונקים, בדרכם הארוכה אלינו.
למשל, הקטניות מייצרות חלבון הקרוי לגמוגלובין (Leghemoglobin), המשמש לקשירת החנקן. הגן האחראי למולקולה זאת דומה בכל לגן של גלובין קדום המשמש את עולם החי מזה 500 מיליון שנה. ממצא זה הביא חוקרים אחרים להשערה כי גן זה עבר מעולם הצומח לעולם החי כטרמפיסט בחיקו של נגיף.
המוות בשרות הגנים
ראיית הברור הטבעי מנקודת המבט של ״אינטרס הגנים״, מתנת בידי החוקרים מכשיר פרשנות חדש על תופעה אוניברסלית, החלה על כל הרב תאים: המוות. כבר הוצעו תאוריות רבות המסבירות את סיבת ההזדקנות והמוות. יש הגורסים, כמו הייפליק, כי כושר התחלקותם של התאים מוגבל למספר סופי וקבוע מראש של חלוקות, לפיכך מרגע לידתנו אנו נושאים בתאינו שעון ביולוגי המונה את פעימות החיים שהוקצו להם. אחרים רואים בהזדקנות ובמוות תוצאה של שגיאות המצטברות והולכות במהלך השכפול של הדנ״א התאי. שגיאות אלו מביאות את התא לייצר חלבונים ואנזימים לקויים, והללו, בהשתתפם בבניית חומרי התא, גורמים לגל שני של טעויות ומקרבים בכך את התא לגבול השגיאה הקטאסטרופלית – כדברי לסלי אורגל. תאוריה אחרת קושרת את תהליך ההזדקנות והמוות במערכת החיסונית; הגוף מפעיל את נוגדניו שלו עד רקמותיו הוא. אחרים קושרים את ההזדקנות בהתרופפות כושרם של התאים השונים לשמור על השוני הספציפי שלהם. לדוגמא, ככל שהשנים נוקפות נעשים תאי הכבד דומים יותר ויותר לתאי הכליה או העור וכיו״ב, משום כך שוב איש יכולים לתפקד כהלכה כתאי כבד. הדנ״א עצמו מאבד מכושר התיקון העצמי, דבר המביא להצטברות שגיאות ולמות התא. כך או כך אנו ניצבים לפני עובדה תמוהה, שקיבלה ביטוי קולע בפי ג׳ורג׳ ויליאמס: לאחר מעשה ניסים של יצירת אורגניזם מורכב מביצית מופרית אחת, אין אורגניזם זה מסוגל לעמוד בדרישות הפשוטות יותר של שמירת הקיים.
ואכן, לנוכח פלאי הפלאים של מבנה האיברים ותיפקודם, הכיצד לא פיתח הבירור הטבעי מנגנון עלומים המבטיח את נצחיות האורגניזם? ואם אכן כל המכאניזמים הביולוגים בטבע נועדו להפצת הגנים, מה לך אינטרס גדול יותר לגנים מאשר מנגנון המבטיח להם כושר שכפול נצחי? שהרי, די היה שתתרחש באורגניזם קדום כלשהו מוטציה אחת המאריכה את חייו, כדי שתזכה לעידודו של הבירור הטבעי (בדומה למוטציה האלטרואיסטית) עם כל דור חדש, עד שהיתה מביאה להופעתם של יצורים בני אלמוות.
וייסמן, הביולוג הגרמני הנודע שחי בראשית מאה זאת, ניסה להראות שההזדקנות היא, למעשה, התאמה חיובית מבחינת האורגניזם, שכן היא מונעת מפרטים קשישים להתחרות בצעירים ובכשירים מהם על המשאבים המוגבלים של הסביבה. רבים אימצו, ויש הממשיכים לאמץ גישה זאת ואף מוסיפים לה חיזוק בשם ״טובת המין״. דהיינו – הורים קשישים עלולים להעביר לצאצאיהם גנים לקויים, שיש בהם כדי לפגוע בחוסנו של המין כולו. אלא שטענות אלו, הנן, כמובן, מעגליות: חיה מאבדת מכשירותה בהזדקנה; היא מזדקנת משום שהיא מאבדת מכשירותה; מסקנה: היא זקנה כיוון שהיא מזדקנת. במילים אחרות, טענות אלו מניחות את המבוקש ואינן עונות על השאלה מדוע החיה המזדקנת מאבדת מכשירותה.
תשובה מעניית יותר מתבססת על האבחנה שהבחין ויסמן בין תאי הגוף לתאי הרבייה. כזכור יצא ויסמן נגד התפישה הלמרקיאנית, בקובעו שהשינויים המתחוללים בתאי הגוף בהשפעת הסביבה, לא מגיעים לתאי הרבייה ולפיכך לא עוברים בתורשה; תנועת המידע בגוף היא חד-סטרית: רק שינויים המתרחשים בתאי הרבייה מגיעים לתאי הגוף ואף מועברים בתורשה לצאצאים.
קביעתו של ויסמן משמעה שהבירור הטבעי מקנה לאבולוציה את כיוונה באמצעות תאי הרבייה ולא באמצעות תאי הגוף. אבל, וזאת צריך להדגיש, עצם הפעולה של הבירור הטבעי נעשית על תאי הגוף.
הבא נחזור לרגע לדוגמא של הכבשים בעלות הצמר הסמיך והדליל. הסיבה ללידתו של טלה בעל צמר סמיך נעוצה בעובדה ששני אללים רצסיבים לצמר סמיך, שנשאו הוריו, חברו יחד ויצרו מצב הומוזיגוטי. אבל העובדה שרק הוא נותר בחיים בקרה הגדולה שנפלה, נובעת מכך שהבירור הטבעי פעל לטובת הטלה אשר תאי גופו הצמיחו צמר סמיך.
מדוגמא זאת אפשר לנסח את הדוקטרינה של ויסמן במילים אלה: תאי הרבייה אחראים להעברת המידע הגנטי מדור לדור, ואילו תפקידם של כל שאר תאי הגוף הוא להגן על תאי הרבייה, להבטיח את השרדותם ולאפשר להם להעביר המידע הכלול בהם לדורות הבאים.
מחלוקת תפקידים זאת עשויה להיווצר ביניהם זיקת גומלין מאוד מעניינת. עקב הסביבה העויינת ומאבק הקיום האכזרי, עתיד האורגניזם, במוקדם או במאוחר, לספות בדרך זאת או אחרת. כיוון שכך, כלום לא יהיה זה בזבוז להשקיע אנרגיה ומשאבים יקרים בפיתוח מנגנונים המבטיחים לאורגניזם חיי נצח – שממילא לא יזכו להתמצות? האם לא יהיה זה נבון ומועיל יותר להשקיע תחת זאת במנגנוני פריון יעילים, המסוגלים להעמיד בקצב מהיר צאצאים רבים ולהבטיח בכך את המשכיות הגנים?
הבה נבחן יותר בפירוט שאלה זאת. כדי לפתח מנגנונים יעילים לתיקון שגיאות הנופלות במהלך ההעתקה והשכפול של הגנים, דרושים, כמובן, אנרגיה ומשאבים לא מעטים. השקעה כזאת מוצדקת בתאי הרבייה, באשר כל שיבוש שם משפיע מיידית על כל הגוף. לא כן הדבר ליתר תאי הגוף, שהרי כל מה שנדרש מהם הוא לתפקד ולהתקיים עד למימוש ההפרייה ולידת הצאצאים. על-פי קו מנחה זה צריך הבירור הטבעי לעודד בגרות מהירה, כושר פריון גבוה ומשך חיים קצר אצל מינים השרויים בסכנה מתמדת, ואילו אצל פרטים השרויים בסביבה בטוחה יותר – חייהם יהיו ארוכים יותר וכושר פריונם נמוך יותר. ואמנם, כך קורה לעכברים ולעטלפים. הראשונים מולידים הרבה צאצאים וחייהם קצרים; השניים, המוגנים יותר ובעלי מנגנון התחמקות משוכלל, חיים יותר זמן. על-פי תפישה זאת, איפוא, העדיף הבירור הטבעי את המנגנון של כושר הרבייה הגבוה והיעיל על-פני המנגנון של חיי נצח. עם זאת, אין בכך כדי להשיב על השאלה מדוע לא פעל22 הבירור הטבעי לדחיית ההזדקנות ע״י העדפת גנים המאריכים את חייו של הפרט. שהרי מבחינת ההשקעה כדאי יותר להשקיע בפרט המסוגל להקים 50 דורות של צאצאים, מאשר בפרט המוליד דור אחד ומת.

לסיר פיטר מדאוור תשובה מעניינת. לדעתו, דחה הבירור הטבעי, הלון ודחה, את תחילת פעולתם של גנים מזיקים, כדי להבטיח את התרבות הפרטים. תהליך ההזדקנות, איפוא, הוא התוצאה הקולקטיבית של גנים מושהים אלה, אשר פעולתם משתחררת בשלב מאוחר של החיים. ומאחר שהם באים לביטוי לאחר שפרט סיים להוליד את מכסת הצאצאים שלו, אין לבירור הטבעי עניין לעשות להכחדתם. למשל, אם אוכלוסיה של פרטים נושאת גנים הגורמים להופעת סרטן בגיל מאוחר, יחמוק גן זה מפעולת הבירור הטבעי ויעבור ללא מפריע בתורשה לדורות הבאים. שכן, מחלת הסרטן תתקוף את הפרטים לאחר שכבר הולידו צאצאים, ולפיכך אין שום יתרון בירורי לנטולי גנים אלה על-פני נושאיהם.
אפשר וזאת הסיבה לסובלנות הרבה שמגלה הבירור הטבעי כלפי תופעת ההשמדה העצמית בקרב העכברים הכיסנאים שהזכרנו בפתח הדברים, המתים משטפי דם פנימיים תיכף להזדווגות. שכן לבירור הטבעי אין עניין בעכבר כפרט, אלא בגנים שלו. הוא הדין בגנים. לדידם העכבר הוא רק אמצעי להפצת שכפוליהם. מבחינתם אין נפקא מינה אם נושא הגנים הוא חיה מלכותית כמו האריה או טפיל עלוב כמו דג החכאי (תיכף ללידתם נצמדים זכרי החכאי לראש הנקבה ופותחים בחיים טפיליים מובהקים: גופם נטמע בבשרה והם מקבלים את מזונם מדמה. במרוצת הזמן מתנוון גופם לכדי שקיק של זרע המשמש לה להפריית הביצים).
פריחת הבמבוק – אחת ל-120 שנה
אם אנו מקבלים קו מחשבה זה, אין מנוס מהמסקנה העגומה כי מה שעומד בינינו לבין חיי אלמוות איננו חוק ביולוגי עליון, נוסח חוק שימור החומר בפיסיקה, אלא תוצאה של חישובי כדאיות ברמת הגנים.
אפשר לשאול, כמובן, אם איננו מייחסים תחכום מופרז למנגנון הפשטני והגס של הבירור הטבעי. שאלה זאת קשה להשיב עליה. מכל מקום, שלל המנגנונים שהתפתחו בטבע למטרות השרדות, הם מגוונים ומתוחכמים להפליא. למשל, מנגנון ההולדה המושהית והסינכרונית.
מעקב אחר זן של במבוק, הנמשך ברציפות למן שנת 999 לספירה, מראה כי זן זה פורח ומייצר זרעים אחת ל-120 שנים. משמע, במשך 12 עשורים נוהג הבמבוק כעשב המתפשט באמצעות חוטרים, ורק בשנה ה-120 הוא פורח, מפרה עצמו, מפיק זרעים ותיכף לכך מת. אך בכך לא תמה סאת הפליאה. גם כאשר שתלו חוטרים של במבוק זה במדינות אחרות, בעלות אקלים שונה, הוא פרח בו-זמנית בשנה ה-120, בהתאם לשעון הפנימי שלו. מעבר לעניין המרתק בטיבו של מנגנון ביולוגי המסוגל למנות במדוייק מניין יוצא דופן כזה של זמן, נשאלת השאלה מה היתרון שיש לפריחה מושהית וסינכרונית זאת לכשירות הדארוויניסטית של הבמבוק. ובאותו הקשר, מה מביא את הנימפות של צרצרים (החיים ביערות של מזרח ארה״ב), לעלות אחת ל-17 שנים ממעבי האדמה, ואז, תוך שבועות ספורים, להתבגר, להטיל ביצים ולמות?
סטפן ג׳י גולד, בספרו Ever Since Darwin, מביא את גרסת האבולוציוניסטים הרואים בלידה המונית ובו-זמנית יתרון גדול למינים השרויים בסביבה23 שורצת טורפים. שכן, אפילו יתכנסו כל הטורפים כדי לאכול בהם בכל פה – לא יוכלו להשמיד את כל האוכלוסיה.
לעומת זאת, כאשר ההתרבות פרושה לאורך זמן רב, יש בכך כדי לספק לטורפים מזון נוח בכל ימות השנה. ואמנם, כמות הזרעים שמפיקה אוכלוסיית הבמבוק בעקבות הפריחה, עשוייה ליצור שטיח בעובי של 15 ס״מ מעל פני הקרקע, כמות שיש בה כדי להשביע את הארי ועוד להותיר קומץ להמשכיות המין.
אצל הצרצרים, המחזוריות בת 17 השנים חושפת עיתוי מופלא ביותר. 17 שנות חייהם בינות לשורשים התת-קרקעיים מהווים פרק זמן ארוך יותר מתוחלת החיים של כל אחד מטורפיהם הטבעיים. מה גם שהוא מספר ראשוני שאינו מתחלק בשום מספר אחר, אלא בעצמו. למה הדבר חשוב? כי אילו היו עולים על פני האדמה אחת ל-15 שנים, ונניח שמחזור החיים של הטורף העיקרי שלהם הוא חמש שנים, הרי שהיו נטרפים ע״י כל דור שלישי של טורפים. לעומת זאת, מאחר שהם עולים אחת ל-17 שנים, הרי שייפגעו ע״י דור חדש של טורפים רק אחת ל-85 שנים (5X17 = 85).
הרצון החופשי מול התכתיב הגנטי
תפישה זאת של הבירור הטבעי המתנהל ברמת הגנים, מעניקה מעמד שווה של אינטרס וכדאיות להתנהגותו של באבון, המפיל עצמו עם פנתר מראש צוק כדי להגן על עדתו, ולפינגווין הדוחף את חברו מראש צוק למים כדי להפיח באחרים אומץ לקפוץ בעקבותיו; לכלב הפרא האפריקני, המקיא את טרפו כדי להאכיל את גוריו, ולאריה ולקוף הלנגור ההורגים את גורי קודמיהם כדי להבטיח האינטרסים שלהם; לצבוע המסוגל לטרוף את גוריו בבולמוס האכילה ולפילה המניקה ברצון את צאצאי אחיותיה ואשר אינה זזה ימים מפילונים חולים עד החלמתם. העמדת הבירור הטבעי על רמת הגנים פירושה שלילת כל אבחנה ערכית וסמלית בין התנהגותו של זכר כנימה, המזדווג עם אחיותיו בחלל בטנה של אמו – בלי שיצא מתחומי גופה כל ימי חייו, לבין דג הסלמון השוחה כ-2,500 ק״מ במעלה הנהר כדי להפרות את ביציה.
פירושה של תפישה זאת – ניתוק הקשר האינטואיטיבי בין הדחף לבין בעל הדחף, עד כדי הצדקת ההזנחה של האם את גורה החלש והעדפתה את החזק מטעמי כדאיות, מצד אחד, והשלמה עם גזר דין זה מצד הגור, שכן במותו יתרום להשרדות 50% מהגנים שלו השוכנים אצל אחיו החזק והמוצלח יותר.
כמובן, ניתן לשאול האם אנו נוהגים בתבונה בכורכנו את כל תופעות האלטרואיזם בחבילה אחת. למשל, התנהגות העש והבאבון. קיימות אוכלוסיות של עש המורכבות מפרטים אכילים ומפרטים בעלי טעם דוחה. אם בירור שאירים אכן תקף, גרס החוקר בלֶסט, צריך אורך חייהם של הפרטים האכילים להיות קצר יותר מהלא אכילים. שכן, ככל שמספרם באוכלוסיה יהיה גדול יותר יתנסו הצפורים בפחות נסיונות מרים, תרתי משמע, ולפיכך ירבו לתקוף את האוכלוסיה בכללותה.
ולהיפך: ככל שמספר הפרטים הלא-אכילים יהיה גדול יותר באוכלוסיה כן יימנעו הצפורים מלנסות לטרוף אותם. ואכן, הממצאים בשטח אשרו את הנחתו של בלסט. המחקר העלה כי הפרטים האכילים נתקפים לעת בגרותם בהתרגשות גדלה והולכת הגורמת להם להתעופף עד אפיסת כוחות, מעוף המסתיים במוות. תוצאותיו של מעוף-התאבדות זה קיצור חיי הפרטים האכילים לעומת הבלתי אכילים. אם-כן, לפנינו התנהגות המוכתבת במלואה ע״י צווים גנטיים, אך האם ניתן לגזור גזרה שווה גם על התנהגותו של הבאבון? כאשר יוצא ראש העדר למאסף ומתייצב מול הנמר האימתני, האם התנהגות זאת צוויה וכפויה עליו עד כדי שלילת כל אלטרנטיבה אחרת, או שמא היא רק מונחית והוא יכול לנהוג פעם כך ופעם אחרת בהתאם לנסיבות ולעוצמת נחישותו?
הגנבת הרצון החופשי כבר ברמה של חיות מצביעה על הבעייתיות הגדולה של הסוציוביולוגיה בכל הקשור להתנהגות האנושית: עד כמה מושפעת התנהגותנו מצווים גנטיים שנרכשו מעולם החי, ומה משקלם של צווים אלה ביחס למורשת התרבותית? במילים אחרות – הצגת החמלה, הרחמים, ההקרבה העצמית, הנכונות לעזור לזולת כפועל יוצא של תכתיב גנטי, משמעה להציג את האדם כיצור דטרמיניסטי ולקעקע את הבסיס הפילוסופי של המוסר, לפיו מותר האדם הוא במאבק המתמיד בין ישותו הרוחנית-מוסרית לבין המורשת הייצרית-חייתית שקיבל מן הטבע. אם, כהצעתו של וילסון, הפסיכולוגיה היא ענף של הסוציוביולוגיה, אזי יש לחפש את המפתח להתנהגות הפסיכית שלנו לא במבנה ובאופן פעילותה של הנפש – אלא בגנים.
כך, למשל, אהבת הורים היא תוצאה של ״הקשר האכזרי״ ושל תחשיבי רווח והפסד ביחס להשקעה, ולא פועל יוצא של רגש ואהבה לשמם.
כאשר מצווה משה את העם להרוג במדיינים, (במדבר ל״א) ״כל זכר בטף וכל אישה יודעת איש למשכב זכר״, אך להותיר בחיים ״כל הטף בנשים אשר לא ידעו משכב זכר״ – אין זה אלא גרסה חמורה של הצו הגנטי של האריה, המורה לו להרוג את גורי קודמיו ולהביא להפלת הלביאות ההרות מזרע לא לו. זאת ועוד, אם האינטליגנציה, השאפתנות, התוקפנות, הנטייה לשלוט, החמלה והאכזריות הן תכונות שוות ערך לטפירים, לניבים ולשרירים, אשר כולם כאחד הם אמצעים שפותחו ע״י הגנים להגברת סיכויי השרדותם ותפוצתם, למה להעדיף את החינוך לחסד ולאהבה על-פני האנוכיות והתוקפנות? ובכלל, איזה ערך או חשיבות יש לשאיפות הטרנסצנדנטליות של האדם, אם הטעם הראשון והעליון לקיומו הוא השרדות הגנים?
כלום לא פתח בכך וילסון, כטענת מתנגדיו, תיבת פנדורה?
הבה נציץ לרגע לתוכה. אם הבירור הטבעי מותיר רק את התכונות הטובות והמועילות לגנים, ואם אנו אכן נושאים בקרבנו גנים לתוקפנות, לאנוכיות, לניאוף וכו’, אזי חלילה לנו לדכא ביטויים טבעיים וחיוביים אלה של הגנים.

האדם איננו בצל
האנתרופולוג דונלד סיימונס, מחבר הספר הפרובוקטיבי The Evolution of Human Sexuality, איתן בדעתו שההבדלים הרגשיים וההתנהגותיים בין גבר לאשה אינם תולדה של חינוך ותרבות, כי אם משקפים24 את השוני הביולוגי-אבולוציוני של טבעם המיני, המוטבע גנטית במוחם ובהורמונים שלהם. מאחר שהזכר פטור מהריון ובכוחו להפרות מספר בלתי מוגבל של נקבות, הוא מופעל הרבה יותר ממנה ע״י גרויים מיניים. לא כן האשה, משעה שהרתה לגבר אחד אין היא יכולה להרות לגברים נוספים ולמיניותה אין שום תפקיד התרבותי עד לאחר הלידה. כיוון שכך היא זקוקה יותר להגנתו ולעזרתו של הגבר. לצורר זה התפתחה במהלך האבולוציה ליצור יותר רגשי ובעל נטייה לקשר יציב וקבוע. זאת ועוד, כדי להבטיח את יציבותו והתמדתו של קשר זה – היא הפכה זמינה מבחינה מינית בכל ימות השנה והמירה את סימני הייחום של נקבות היונקים (המעידים על זמינותן המינית) בעור חלק ומתוח שתפקידו לגרות את חושיו של הזכר האנושי ולבשרו על כושר פריונה. סיימונס גורס כי לעובדה שגברים בגיל העמידה מסוגלים לעורר ענין מיני בנשים צעירות – הרבה יותר מאשר נשים מבוגרות בגברים צעירים – יש שורשים ביולוגיים אבולוציוניים: גבר בגיל העמידה אינו עובר בלות מינית ובכוחו להוליד צאצאים בריאים וחסונים. לא כך האשה, לפיכך קמטיה עשויים להעיד על אבדן כושר פריונה. זאת ועוד, סיימונס מעורר את זעמם של הפמיניסטיות ושל חסידי התיזה הסביבתית בהציעו שורשים ביולוגיים לשורה שלמה של איפיונים גבריים נוספים, אשר נוטים כרגיל לייחסם לתרבות. למשל, הגברים קנאים יותר מאשר הנשים משום שהזכר לעולם אינו בטוח באבהותו על צאצאיו, בעוד הנקבה פטורה, בכל מקרה, מהטלת ספק באמהותה על צאצאיה. האבולוציה, איפוא, פיתחה את קנאת הגבר כמנגנון שנועד להגן על השקעה שהשקיע בקשר עם הנקבה והבא למנוע ממנו ליפול בפח הקוקייה. גם לנטייה הגברית לריבוי יחסים עם נשים יש תשתית סוציוביולוגית, שכן העובדה שהזכר יכול להפרות מספר בלתי מוגבל של נקבות ושהאינטרס של הגנים הוא להשיג תפוצה מירבית, יצרה לחץ של הבירור הטבעי לכיוון של נטייה פוליגמית אצל הזכרים ולפיתוח מערכת גרוי מינית המופעלת ע״י תחלופה של נקבות. לדוגמא, כאשר מותירים פר עם נקבה אחת, הוא יזדווג איתה מספר פעמים, אך עד מהרה יאבד את הענין בה ולא יחדשו גם אם ירחיקו אותה ויחזירוה אליו לאחר זמן. לעומת זאת, אם יכניסו למכלאתו פרות חדשות בזו אחר זו הוא יזדווג עם כולן באותה להיטות וללא סימני עייפות.
אך נחזור לוילסון. עמדתו אמנם מתונה הרבה יותר, אך רבים חוששים כי די בהנחת ההתנהגות המוסרית על בסיס גנטי, ולו באופן חלקי, כדי לחדש את ההכשר הנתעב לתורות גזעניות. ואכן, למן פרסום ספרו של וילסון נטוש פולמוס חריף סביב הסכנות שבהנחת דטרמיניזם ביולוגי, מכל דרגה שהיא, ביסוד ההתנהגות האנושית; במקום, ואפילו לצד, הפרימט התרבותי. אומר על כך רוברט מרטין: ״האדם איננו בצל שאפשר לקלף ממנו את הקליפות החיצוניות של התרבות עד לחשיפת הגלעין הביולוגי״. לדעת מרטין, כל נסיון לנתח את החלק הביולוגי של ההתנהגות האנושית, בלי לחדור לניתוח החלק הנובע מהתרבות האנושית – נדון לכשלון. גם לסיר ג׳וליאן הקסלי, מבכירי האבולוציוניסטים של תקופתנו, לא היו ספקות בנושא זה. בספרו Evolution in Action מצביע הקסלי על זרם התרבות כעל יחידת אבולוציה עיקרית בהתפתחות האדם. השפעת הגורם הגנטי בעיצוב המין האנושי, גרס הקסלי, היא משנית בהשוואה לאמנויות, לקודים המוסריים, לאמונות הדתיות -ובראש וראשונה לידע ולאידאות.
בשל ההתקפות החריפות על ספרו ועל המשמעויות החבויות בו, נדרש וילסון שוב ושוב להבהרת עמדתו, בראיון לניו-יורק טיימס, מיום ה-9 בנובמבר 1975, חיווה וילסון דעתו כי רק ל-10% מההתנהגות האנושית יש בסיס גנטי, ואילו היתרה נובעת מהשפעות סביבתיות: חינוך, חברה, תרבות וכיו״ב. אלא שגם עשרה אחוזים אלה מעוררים את חמתם של חסידי הסביבה, הרואים בתרבות את היורש הבלבדי של הביולוגיה בעיצוב האדם והחברה האנושית. אך וילסון בשלו. יתירה מזאת, הוא משוכנע כי לא ירחק יום ונוכל לאתר גנים ספציפיים האחראיים לחלק מהתנהגויותינו. ככלות הכל, מדגיש וילסון, מרבית האבולוציה הגנטית אשר עיצבה את התנהגותו החברתית של האדם, התרחשה בחמישה מיליון השנים שקדמו להופעת התרבות. זאת ועוד, בחלקה הארי התפתחה התרבות ב-10,000 השנים האחרונות – עם הופעת החקלאות והחיים האורבניים, פרק זמן זה הוא, לכל הדעות, קצר מכדי להשפיע באופן גנטי על הטבע האנושי. אלמלא היה זה כך, היו צריכים להיות הבדלים גנטיים בין יוצאי חברות ציידים לבין יוצאי חברות חקלאיות.

המרחק האינדיבידואלי של בני אדם וחשך הורים אצל קופים
בדבר אחד לבטח אין מחלוקת, בדימיון שבין התנהגויות ותכונות מסויימות של בעלי חיים לאלה של בני-אדם, אפילו נתרחק מכל נטייה אנתרופומורפיסטית (נטייה להאנשה). ניקח, למשל, שתי דוגמאות משני קצות הטווח ההתנהגותי:
המרחק האינדיבידואלי של בעלי חיים והשפעת חשך הורים על צאצאים.
המרחק האינדיבידואלי של בעלי חיים הוא המרחק המינימלי אשר מרבית בעלי החיים מקפידים לקיימו בינם לבין פרטים אחרים מבני מינם. מרחק המהווה פשרה בין הקירבה ההדדית הנדרשת לפרטים חברתיים, לבין הריחוק ההכרחי להפגת האיום שבקירבה יתירה. לכל מין כמעט מרחק אינדיבידואלי אופייני (ניתן למודדו במדוייק בהמצא הפרטים מחוץ לטריטוריה שלהם). למשל: אצל הדרור – 15 ס״מ, שחף שחור הראש – 30 ס״מ, הפלמינגו – 60 ס״מ, העגור – 175 ס״מ וכו’. אצל האדם שונה המרחק האינדיבידואלי מתרבות לתרבות. על-פי מחקרו של אדוורד הול, העמים הים-תיכוניים, למשל, סובלניים יותר לצפיפות ולקירבה גופנית (בעת שיחה או במסעדה) מאשר העמים הצפון-אירופיים. כתוצאה מכך, אנגלי הנפגש לשיחה עם איטלקי רואה בעמידתו הקרובה מדי אליו אות לגסות רוח, בעוד שהאיטלקי מפרש את עמידתו המרוחקת של האנגלי כחוסר נימוס וקרירות. בולטים בנטייתם הקיצונית לפרטיות הגרמנים. מטעם זה, סבור הול, בתי מלון ובתים פרטיים רבים בגרמניה מצויידים בדלתות כפולות להבטחת פרטיות היושבים מאחוריהם. אגב, מחקר שנערך לפני 10 שנים לערך בארה״ב, בקרב אסירים, הראה כי המרחק האינדיבידואלי של אסירים אלימים, שמתחת לו הם מגיבים בתוקפנות, הוא מטר אחד. עורך המחקר, אוגוסטוס קינדל, סבור שאפשר להשתמש בניסוי זה כדי לעמוד על רמת האלימות של האסירים, ואף להסתמך על המרחק האינדיבידואלי כדי לשחרר בערבות את הפחות אלימים.25 הדוגמא השניה, כאמור, היא השפעת חשך הורים על צאצאיהם. בנסיונו המפורסם של הרי הרלוב, בודדו תינוקות של קופי רזוס מאמהותיהם לפרקי זמן של 3, 6, ו-12 חודשים. לאחר שהושמו בכלובים מבודדים הציבו בהם דחלילי קופים עשויים מסמרטוטים והתינוקות התרפקו עליהם כאלו היו אמהותיהם. בתום תקופת הבידוד השיבו אותם לחברת קופים בני גילם אך הם הפגינו התנהגות חריגה, אשר בחברה אנושית היו מגדירים אותה כפסיכוטית: עודף פעילות, מצד אחד, וישיבה משמימה באפס מעשה ובעיניים בוהות בחלל, מצד שני, כיצורים אוטיסטים לכל דבר. כן הרבו לבכות ולמצוץ אצבעות ידיהם ורגליהם. התברר גם שבידודם שיבש כליל ובאורח קבע (אצל אלה ששהו שנה בבידוד) את פעילותם המינית. הזכרים לא ידעו כיצד להזדווג עם הנקבות, ואילו האחרונות דחו בתוקף כל התקרבות מצד הזכרים. באותם מקרים שנאנסו ע״י זכרים מנוסים וילדו, זנחו את תינוקיהן ודחו אותם מעליהן בגסות כל אימת שביקשו לינוק.

גמישות וקשיחות בשימוש בכלים
שתי דוגמאות אלו מחייבות זהירות מרובה בגין ההקבלה החיצונית שבינן לבין התנהגויות אנושיות. הטבע מספק רפרטואר של התנהגויות דומות, חלקן מורכבות להפליא – כמו גני הפטריות ממוזגי האויר שמטפחים הטרמיטים במעבי תיליהם – וחלקן נתפשות על-ידינו כבעלות מטען רגשי עז, כמו הקשר המונוגמי המקובל על 92% ממיני העופות והמאופיין ע״י מסירות ונאמנות מעוררות השתאות. ההבדל הגדול הוא שדפוסי התנהגות חברתיים אלו מועברים מדור לדור באמצעות הגנים ומוכתבים בקשיחות ע״י צווים גנטיים. המבנה המקורי המורכב להפליא של הטרמיטים המבטיח לגני הפטריות טמפרטורה יציבה של 30 מעלות צלזיוס, נעדר כל גמישות. די בשיבוש התנאים הסביבתיים או בשינוי הפרשת אותות הזהוי או בהפרת הקודים ההתנהגותיים כרי להביא כליה גמורה על כל המושבה ואולי גם על המין כולו. ההתנהגות האנושית, לעומת זאת, מבוססת בחלק זה או אחר על העברת מידע ע״י הוראה ולמידה. זאת ועוד, היא ניזונה מרפרטואר גמיש ובעל מגוון עצום של בחירות וברירות המאפשרות לפרט להתאים עצמו לנסיבות ולתנאים המשתנים של הסביבה, תוך הפעלת תבונה וחוכמה.
ניקח, למשל, השימוש הנעשה בכלים ברמות השונות של החיים בטבע. מקובל למנות את שושלת האדם מיצור קדמון, ספק קוף ספק אדם, הומו הביליס, שהיה הראשון לעשות שימוש בכלים שיצר לצרכיו. אך שימוש בכלים נעשה הרבה זמן לפניו, ע״י בעלי חיים נחותים ופשוטים ממנו לעין שעור.
מינים אחרים ממשפחת הצרעות נוהגים לסגור את הכניסה לקיניהם ע״י אבנים קטנות; הארי-נמל המכונף צד נמלים ע״י יריית חול בראשן. בקרב העופות, עושה מין של פרוש מאיי הגלאפגוס שימוש בקוץ שהוא מחזיק במקורו כדי לדוג חרקים מתוף הסדקים שבקליפות עצים; הפרס המצרי נוטל אבנים מהקרקע ומטילן מהאויר על ביצי יענים כדי לשבור את קליפתן הקשה. הלוטרה הימית שוברת צדפות באמצעות אבנים שהיא מניחה על כרסה. צפור המרבו שולפת במקורה את חיפושיות הזבל מצואת הפילים, שוטפת אותן במים ורק לאחר שהן רחוצות ונקיות, היא בולעת אותן. המאפיין שימושים אלה הוא שהם אינם פרי של חשיבה אינטליגנטית ועפי״ר אפילו לא של למידה. זוהי התנהגות גנטית שהתפתחה במרוצת האבולוציה מתור דפוס התנהגותי דומה, אשר מילא פונקציה מסויימת אצל אבותיהם. לעומת זאת, השימוש בכלים אצל הקופים הוא פרי חקוי ולמידה.
במשך שנים שימשו להקות של קופי מאקק שבאיי קושימה, נושא למעקב מדוקדק. ב-1952 פיזרו המדענים תפוחי אדמה מתוקים על החוף כדי להעשיר את התפריט של הקופים. לא חלפה שנה ונקבת מאקק, המכונה אימו, נטלה תפוח אדמה לים ושטפה במים את החול שדבק בו. תוך חמש שנים אומץ מנהג זה ע״י 80% מאוכלוסיית הקופים (הקופים הקשישים וכ-80% מהנקבות מאנו להסתגל להגיינה החדשה).
אגב, אותה קופה מחוננת, אימו, המציאה ב-1955 טכניקה גאונית חדשה לאיסוף מזון. לצד תפוחי האדמה נהגו המדענים לפזר על החוף גם גרעיני חיטה, אותם נהגו הקופים ללקט בעמל רב ומבין גרגירי החול. יגיעה זאת לא מצאה חן בעיני אימו. בהברקה נדירה של ״האורקה״, נטלה מלוא חופניים חול מעורב בגרגירי חיטה והשקיעה תוכנם במים: החול שקע מייד ואילו את גרגירי החיטה שנותרו צפים במים אספה והכניסה לפיה. לא נקפו ימים רבים וכל הלהקה, להוציא הזקנים השמרנים הלכו בעקבות אימו. השימוש בכלים רווח במידה לא מעטה גם בקרב השימפנזות: הטלת מקלות ואבנים באויביהם (אף כי באחוז פגיעה נמוך של 10% לערך), שימוש בקני עשב ללכידת נמלים בתוך סדקי עצים, שימוש במקלות כבמנופים לפתיחת מכסים כבדים של ארגזים, שימוש בקוצים לניקוי שיני הקופים הצעירים ועוד.
עורות חיים קפואות ומתפתחות, ושגיאות גורליות של האבולוציה
אם, כפי שדוגמאות אלו מלמדות, מסוגלים קופים לעשות שימוש נבון ומגוון בכלים ואף להוריש ידע זה בלמידה, מדוע לא פיתחו ושיכללו מיומנות זאת במרוצת מיליוני שנות קיומם?
התשובה טמונה בגידול מוחי מדהים ויחיד מסוגו בטבע, שראשיתו לפני 3 מיליון שנה לערך אשר ליווה את התפתחותם של מינים אחרים של הומינידים (אדם-קוף), גידול שפסח על הקופים. מקובל כיום לראות בדריופיתקוס, קוף קטן שחי לפני 20 מיליון שנה, את האב הקדמון המשותף של קופי האדם החיים עמנו כיום ושל האדם. מעריכים כי לפני 14 מיליון שנה התפצלו צאצאיו של הדריופיתקוס לשלוש שושלות נפרדות: שושלת קופי ענק, ג’יגנטרופוס, שנכחדה כליל; שושלת קופי-האדם שהוציאה מתוכה את הגורילות, השימפנזות והאורנגוטנגים של ימינו; והראשון לשושלת ההומינידים רמאפיתקוס, אדם-קוף קטן אשר נטש את היער ספק על שתיים ספק על ארבע כדי ללקוט זרעים ושורשים בסוואנה הפתוחה.
אין זאת שמעשיו של רמאפיתקוס צלחו בידיו, שכן שרידיו נמצאו באירופה, בהודו, בפקיסטן ובסין. גם שושלת ההומינידים, שיצאה מחלציו עשתה חיל. הראשון בהם, האוסטרלופיתקוס אפריקנוס, הופיע על במת העולם לפני 5 מיליון שנה לערך. הוא היה יצור קטן למדי, אך בעל נפח מוח של 450 סמ״ק – כנפח מוחן של הגורילות והשימפנזות בעת ימינו. גם בצורתו דמה להן ואעפי״כ היה שונה: הוא הילך כמעט זקוף ולגולגלתו ולשיניו היו איפיונים אנושיים מובהקים. תוך 3 מיליון שנה פינה מקומו ליורש חזק ואינטליגנטי גדול ממנו, האוסטרלופיתקוס רובוסטוס. אלא שבכך הגיעה שושלת האוסטרלופיתקוס לסוף דרכה. איש אינו יודע מדוע. אולי משום שהשינויים שהתברך בהם קפאו על שמריהם או שהתפתחו בכיוון לא נכון, או שלא היה בהם די לעמידה מוצלחת במאבק הקיום הקשה ותנאי הסביבה העויינים של הסוואנה. כך או כך תרומתו של האוסטרלופיתקוס לאבולוציה הסתכמה במספר מאובנים נאים שהותיר ותו-לא.5
לכאורה אפשר לראות במשך חיים בן 5 מיליוני שנים סיפור הצלחה. ככלות הכל, ההומו ספיאנס נולד לפני 100,000 שנה ונכחד תוך כמה עשרות אלפי שנים ואילו אבותינו הישירים – ההומו ספיאנס-ספיאנס – דרכו על במת העולם רק לפני 40,000 שנה, ואין שום26 ודאות שיסיימו את אלף השנים הבאות. אך בקנה המידה האבולוציוני נמדדת ההצלחה בשתי אמות מידה: מידה של זמן ומידה של השתפרות, או בניסוחו של ז’ואן הקסלי: ״פרוגרס ביולוגי״, שמשמעו שיפור שאינו חוסם דרכם של שיפורים נוספים. כל פרוגרס, קובע הקסלי, נעשה בסדרה של שיפורים ובמסגרת כיוון כללי, עד שהוא מגיע לנקודת עצירה. לפעמים נשאר המין תקוע בנקודה זאת מיליוני שנים עד להכחדתו ולפעמים הוא יוצא מקפאון זה ע״י סטייה מפתיעה לכיוון חדש. צורה קפואה כזאת, אשר רק נציגים ספורים ממנה חיים עמנו כיום, היא קונכיית-ים קטנה, הלינגולה, אשר לא השתנתה זה 400 מיליון שנה. כזהו גם דג הראה האוסטרלי, הזהה לאבותיו המאובנים מלפני 150 מיליון שנה. סוג אחר של צורה מתמידה, השונה מה״מאובנים החיים״ בכך שהקבוצה בשלמותה מוסיפה להתקיים ולשגשג, היא קבוצת הנמלים, אשר לא חל בה שינוי משמעותי זה 50 מיליון שנה. אגב, גם בצפורים, כמכונות תעופה, לא חלו שינויים של ממש ב- 20 מיליון השנים האחרונות. הסיבה לקפאון או להתמדה זאת היא שבמהלך האבולוציה הגיעו צורתם ותכונותיהם להתאמה ולאיזון כה מושלמים עם התנאים השוררים בגומחה הספציפית שהם תופשים בטבע, עד כי כל מוטציה שמתחוללת בהם אין בה, מבחינת הבירור הטבעי, שום יתרון ולפיכך היא נכחדת.
אבל, לא תמיד קוצרת האבולוציה תוצאות מוצלחות כל כך. יש והיא פותחת את מסעה במה שמסתבר בדיעבד כטעות קטנה וכמו-שולית, ואשר תוצאותיה הגורליות באות לביטוי רק כעבור עשרות מיליוני שנים.
כך ארע, למשל, עם החרקים. החרקים מעבירים את החמצן לרקמותיהם – הישר מן האוויר, באמצעות מערכת עדינה של צנוריות. זוהי שיטה יעילה וחסכונית יותר מראות, כל עוד שומר החרק על מימדי גוף קטנים. אך אילו גדל למידתו של עכבר לא היה יכול להתקיים.
מסקנה; האבולוציה והבירור הטבעי, אשר ניתבו את החרקים למערכת זאת של נשימה, חרצו בכך לא רק את גודלו המירבי, כי אם, חשוב הרבה יותר, גזרו עליו לחיות עד קץ הימים בהגבלה שכלית, שכן מידות גופו הזעירות אינן מאפשרות לו לגדל מוח גדול. ואולי טוב הדבר, מעיר הקסלי, כי אילו פיתח החרק אינטליגנציה גבוהה, הוא היה כיום השליט הבלבדי על העולם במקומנו.
עניין זה של המוח מזכיר טעות נוספת של האבולוציה, אולי הרת גורל יותר מקודמתה, שארעה אצל מפריקי-רגל. אצל בניה של מערכה זאת, הכוללת את החרקים למיניהם, העכבישים, הסרטנים ועוד, בנוי מוחם סביב הוושט. במהלך האבולוציה, ככל שהמוח התפתח וגדל, הלך והתכווץ הוושט בלחץ תאי העצבים שגדלו סביבו. אצל העכבישים והסרטנים הגיעו הדברים למצב כזה ששוב אין הם יכולים לבלוע אוכל מוצק אלא נוזלים בלבד. באופן ציורי אפשר לומר, שבמהלך האבולוציה של העקרבים עמד הבירור הטבעי לפני דילמה קשה: האם לפתח עקרבים טפשים, אך מסוגלים ליהנות מארוחה טובה; או האם לפתח עקרבים המסוגלים לצוד טרף בתבונה רבה, אך לא מסוגלים לבלעו. הבירור הטבעי עשה פשרה בין השניים, אף בשל טעות ארכיטקטונית מטופשת שלו נידונו מפריקי הרגל להגבלה שכלית, ללא כל סיכוי לשינוי כלשהו בעתיד.
אך נחזור לצורות המתמידות שהזכרנו קודם. כנגד הצדפות, הקונכיות והנמלים, המתמידים בצורתם ללא שינוי, מהווה ההתפתחות שעבר הסוס דוגמא קלאסית לפרוגרס ביולוגי רצוף שיפורים.
אביו הקדמון של הסוס בן ימינו, אשר חי ביערות לפני 55 מיליון שנה, היה קטן גוף. גובהו לא עלה על 30 ס״מ והיו לו ארבע אצבעות בכל רגל. ב-15 מיליון בשנים הבאות יצאו מחלציו של אב קדמון זה מינים מגוונים של סוסים, שרובם נכחדו על שושלותיהם. לפני 35 מיליון שנה לערך החל תהליך חדש שבמהלכו נעלמה אצבע אחת מרגליהם, ואילו המרכזית מבין השלוש הנותרות החלה לגדול. תהליך זה קיבל תנופה כעבור 10 מיליון שנה, בקרב אותם מינים שנטשו את היער ועברו לחיות בערבות העשב הפתוחות. ואז, לפני כ-10 מיליון שנים לערך, ארע השינוי הסופי, אשר התייצב לפני כשלושה מיליון שנה כאקוס – אביו הישיר של הסוס המודרני: הבוהן הפכה פרסה ושתי האצבעות האחרות נעלמו כליל. בכך הגיע הפרוגרס הביולוגי לסוף דרכו. שכן הפרסה מייצגת את דרגת ההתמחות הגבוהה ביותר (בתנאים הקיימים). במצב זה לא ניצב לפני הבירור הטבעי שינוי אפשרי הצופן בחובו שיפור ביחס למצב הנוכחי, לא באשר לפרסה או לגודל הגוף או למבנה השיניים הטוחנות. כל אשר נותר לסוס לעשות הוא להתמיד בצורתו הנוכחית.

המוח האנושי – התעלומה הגדולה של הטבע
אם-כן, האוסטרלופיתקוס שלנו שרד יפה במשך מיליונים אחדים של שנים, אף בסופו של דבר נפל בין שני כסאות: הוא לא הגיע לדרגת ההתמחות הגבוהה של הקופים המאפשרת להם להתקיים כבר יותר מעשרה מיליון שנים. מצד שני, השינויים שחלו בו ואשר הציבו אותו על מסלול ההומינידים, קפאו תחתם, משל מיצו את ״הפוטנציאל האבולוציוני״ שהיה גלום בהם. אין זאת שבהגיע שעת המבחן, לא הצליח האוסטרלופיתקוס להסתגל במהירות הדרושה לשינויים בסביבה ולהתאים עצמו לתנאיה. ואולי לא עמד לו כוחו להתחרות ביצור חדש, מיומן ומוצלח יותר, שזה עתה הופיע: הומו הביליס, הראשון בשושלת ההומו. הומו הביליס, כמו האוסטרלופיתקוס רובוסטוס, הוא נכדו של רמאפיתקוס. אביו, אוסטרלופיתקוס אפרנזיס, התפתח ככל הנראה במקביל לאוסטרלופיתקוס אפריקנוס אף כי שרידיו הקדומים ביותר הם בני 3.5 מיליון שנה בלבד. אך בעוד האפריקנוס ורובוסטוס צעדו לתוך מבוי סתום, יצא מחלציו של אפרנזיס, לפני 2 מיליון שנה לערך, הומו הביליס. להומו הביליס היה נפח מוח של 750-600 סמ״ק והוא, אולי, היצור הראשון בטבע שיכל לחזות את צרכיו מכלי מסויים וליצור בהתאם לכך את הכלי הדרוש לו. הומו הביליס ביצר לו עמדה מרכזית על בימת ההומינידים במשך מיליון שנה, עד שפינה מקומו לצאצא זקוף קומה ובעל נפח מוח אדיר של 1100-880 סמ״ק. היה זה הומו ארקטוס, אביו הקדמון של הומו ספיאנס. המוח הוסיף לגדול בהתמדה והגיע לפני 100,000 שנה, אצל אחד מצאצאיו הכושלים, האדם הנאנדרטלי, לנפח ממוצע של 1500 סמ״ק. אלא שמוח בלבד אין בו די. עובדה, האדם הנאנדרטלי נכחד, ואת מקומו תפש לפני 40,000 שנה לערך, ההומו ספיאנס-ספיאנס, בעל נפח מוח של 1450 סמ״ק, אשר מחלציו התפתחה החברה האנושית בת ימינו.
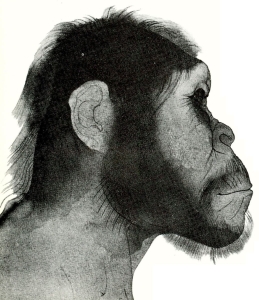
הנה כי-כן, במשך 10 מיליון שנה לערך גדל מוחם של ההומינידים באיטיות רבה יותר. והנה, תוך תקופה קצרה יחסית של 3 מיליון שנה, גדל נפח מוחם ב-300 אחוזים. גידול מהיר ומדהים זה הוא אחת התעלומות הגדולות בטבע. נסיונות חוזרים ונשנים להסביר פנומן זה ע״י המעבר מהיער לסוואנה הפתוחה, ההליכה הזקופה, המעבר מאורח חיים צמחוני לבשרי וכיו״ב, הם במקרה הטוב – דחוקים. גם קופי מקאק מסוגלים להלך מרחקים גדולים על שתי רגליים ולשאת אגב כף מזון בידיהם, ובכל זאת לא חולל בהם שחרור הידיים מהפך מוחי וחברתי כאצל ההומינידים. מה שברור הוא כי התפתחות זאת נשאה יתרון אבולוציוני אדיר ויחיד במינו. שכן מאוכלוסיה בת כמה אלפי פריטים לפני מיליון שנה, מונה היום החברה האנושית יותר מארבעה מיליארד בני-אדם – יותר מכל יונק אחר עלי אדמות.27
האדם – הפאציפיסטי מבין היונקים
בראשית ההתפתחות ההומינידית, כותב וילסון בספרו, היו הלחצים הסביבתיים גורמי התנופה המרכזיים של התפתחותו – כבכל מין אחר של בעלי-חיים. אולם, עם המעבר לחיים בשטח פתוח ולאורח חיים של ציידים, נכנס לפעולה גורם אבולוציוני חדש, פנימי, שהפך במרוצת הזמן בסיס לראשיתה של התרבות. מה שקרה הוא, אומר וילסון, שהשינוי השכלי והחברתי הפך יותר ויותר תלוי ברה-ארגון פנימי, ופחות ופחות נסמך על תגובות ישירות לארועים של הסביבה. ומשעה שהדגש עבר מהחוץ אל הפנים, קיבלה האבולוציה החברתית את כח ההנע העצמי שלה, אשר זכה לתנופה אדירה לפני 40,000 שנה – עם הופעת ההומו ספיאנס-ספיאנס. לפני 10,000 שנה לערך, פותחה החקלאות, דבר שהאיץ את קצב הגידול של האוכלוסיה בשעור מדהים. חברות הציידים הפרימיטיביות פינו דרך להקמת שבטים, נסיכויות ולבסוף מדינות.
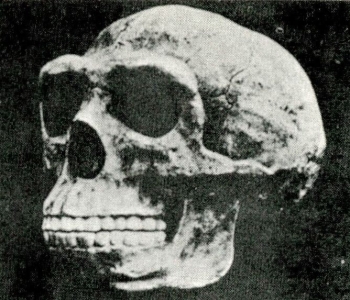
אחד ממאפייני החברה האנושית הוא האלטרואיזם ההדדי, המסועף להפליא והמסוגל ללבוש ולפשוט צורות משעה לשעה. וילסון מציין כי תכונות אלה של שתפנות ואלטרואיזם, העומדות ביסוד מבני החיים החברתיים בטבע, נמצאות בירידה ככל שעולים במעלה הסולם האבולוציוני: מושבות של נבובים, החרקים החברתיים, היונקים והאדם.
במושבות של יצורים ימיים מסדרת הסיפונופורה, למשל, כל הפרטים זהים מבחינה גנטית, אך הם ממלאים תפקידים שונים: חלקם מספקים לגוף קולקטיבי זה את התנועה, חלקם את ההגנה, חלקם את כושר הרבייה וחלקם מהווים את הקיבה של המושבה כולה. כלומר, טרף שנלכד מתעכל בקיבותיהם ומועבר לפרטים האחרים במושבה. במילים אחרות, אפשר לראות בכל אחת מקבוצות פרטים אלו איבר של אורגניזם אחד. אצל החרקים החברתיים השתפנות באה לביטוי בחלוקה למעמדות של התמחות. עם זאת, כל פרט הוא אורגניזם עצמאי ושלם. יתירה מזאת, חיי השיתוף מופרים מפעם לפעם ע״י קונפליקטים ומעשי אלימות – אם כלפי המלכה, או מצד המלכה עצמה כלפי מתחרות אפשריות, ואם מצד הפועלות כלפי פרטים אחרים במושבה. אלא שמעשים אלה הם כאין וכאפס לעומת התוקפנות וניגודי האינטרסים המאפיינים את חברות היונקים. שוב אין אנו פוגשים שתפנות עליונה נוסח החלוקה המעמדית של החרקים. כל חבר באוכלוסיה הוא פרט בודד לעצמו. למעשה, האנוכיות והדאגה לאינטרסים העצמיים שולטות ככיפה ומכתיבות את מערכת היחסים בחברה.
והנה, בהגיענו לאדם, נשברת מגמה זאת. אף שבני האדם הם חולייתנים במבנה החברתי שלהם, כותב וילסון, הם יצרו דפוסים חברתיים בעלי מורכבות מיוחדת במינה. יתירה מזאת, הם שברו את הסייגים של עולם החולייתנים לא ע״י צימצום האנוכיות, כי אם בכוח התבונה שבהם להיוועץ בעבר ולכלכל את העתיד. בכוחם לקיים הסכמים לטווח ארוך ולקבל על עצמם התחייבויות של אלטרואיזם הדדי למשך דורות. ככל שהשתפנות בקרב הקופים היא נדירה, להוציא ביטויים בסיסיים אצל השימפנזות, הנה באדם היא מהמאפיינים החברתיים הבולטים. כתוצאה מכך, רק בחברה האנושית קיימת כלכלה, שכן רק בזכות האינטליגנציה הגבוהה של האדם יכול סחר חליפין, במובן המלא של המילה, להתפתח.

רצח וקניבליזם, מעשים של יומיום בקרב היונקים, הם נדירים לאין שעור אצל האדם. יתירה מזאת, אחד המיתוסים הרווחים, לפיו האדם הוא הטורף היחידי בטבע שהורג לא רק כדי להשביע את רעבונו, איננו מדוייק. אריות וצבועים, למשל, הורגים לפעמים ללא סיבה ברורה. מכל מקום העדויות הרבות שהצטברו בשנים האחרונות על התנהגויות רצחניות וקניבליות אצל היונקים, מפריכות את מסקנתו המפורסמת של קונרד לורנץ (בספרו ״על התוקפנות״), לפיה פציעות ומוות בעקבות עימות כוח על טריטוריה ומאבקי מין, הם תוצאה מקרית ולא מטרה – אין לה סימוכין בטבע. ההיפך הוא נכון, השמדת היריב היא נורמה הרבה יותר מקובלת בקרב חולייתנים שונים מאשר באדם. אילו צפה זאולוג ממאדים באדם והיה משווה התנהגותו למינים אחרים על-פי יחס התקיפות והרציחות לגודל האוכלוסיה וליחידת זמן, היה מגיע למסקנה שהוא נמנה עם היונקים היותר פאציפיסטיים עלי אדמות.
הסיבה העיקרית היא שכל בעלי החיים, להוציא האדם, הם אופורטוניסטים מושלמים. כלומר, התנהגותם אינה מודרכת ע״י מעצורים ונורמות תרבותיים, כי אם ע״י עקרון חד ופשוט: מה טוב יותר עבורם. ואם בנסיבות מסויימות נושא הקניבליזם יתרון בירורי והשרדותי מובהק, כל אוכלוסיית המין תהפוך עד מהרה קניבליסטית.
חרף הבדל מהותי זה בין האדם לשאר בעלי החיים, ואף שהגנים העבירו לתרבות את התפקיד המרכזי בעיצוב התכונות האנושיות, וילסון גורס שהם מחזיקים עדיין במידה מסויימת של השפעה לא רק על תכונות, דחפים, יצרים ונטיות, כי אם גם, במידת-מה, על עיצוב התרבות. כאשר נוצרו חברות האדם הראשונות איפשר להם כושרם השכלי להעדיף את היתרונות או החסרונות שבשיתוף פעולה עם בני קבוצה שכנה של הומינידים. קבוצה אשר גירשה את שכניה וירשה את אדמתם ונכסיהם, שיפרה את מצבה והגדילה את הכשירות הדארוויניסטית של חבריה. משמע, הצלחת המעשה תרמה להפצת הגנים החסונים והאלימים של המנצחים, ובו בזמן נרשמה בזכרון של הקבוצה ושימשה דגם חיקוי ליצירת תרבות של אלימות. במילים אחרות, עקרון הבירור הטבעי חל לא רק על הגנים, כי אם גם על התרבות: דפוסים תרבותיים יעילים ומוצלחים יותר אומצו והתפשטו, ואילו הפחות מוצלחים עברו מן העולם או התמעטו. כך או כך, הזיקה ההדדית בין גנים לתרבות קיימת למן שחר התרבות.
לדעת וילסון, הבנתנו המוגבלת את מוח האדם אינה מאפשרת לנו לדעת אלו תכונות ערכיות קשורות באופן גנטי לתכונות ראשוניות והרסניות יותר. למשל, שתפנות כלפי בני הקבוצה עשוייה להיות קשורה בתוקפנות כלפי זרים, יצירתיות עשוייה להיות הפן השני של נטייה לקניין ולשליטה, להיטות אתלטית צמודה אולי לתגובה אלימה וכו’. כדי להבטיח את המשכיות המין האנושי לאורך זמן, חובה עלינו לחתור לידע טוטלי, עד לרמות של הנוירון ושל הגן. כאשר נגיע לנקודה שבה נוכל להסביר את עצמנו במונחים מכאניסטים אלה, סבור וילסון, נדע שהאנושות הגיעה למצב אקולוגי יציב. תחליף ההפנמה של האבולוציה החברתית, שהחל בקבוצות הפרימיטיביות של האדם הקדמון, יגיע, פחות או יותר, לסיומו. למצב זה, אומר וילסון, תגיע החברה האנושית במאה ה-21. אבל, הוא28 29 מוסיף ואומר, בצטטו את קאמי, עולם מוסבר הוא עולם מוכר. מצד שני, בעולם המשולל אשליות והארות, אדם חש עצמו זר ומנוכר.
וילסון מסכים עם הערכתו של קאמי, אך הוא מנחם עצמו ואותנו בכך שלרשותנו עדיין מאה שנים של עולם בלתי מוכר ומפורש.
הסבר כפול לשירת הצפורים ולמות האסקימוסים
וילסון, אם-כן, מאמין שניתן לאתר את המרכיבים הגנטיים של ההתנהגות הפסיכולוגית-חברתית של האדם ע״י זהוי המרכיבים ההסתגלותיים שלה. למשל, אם ניתן להראות שבנסיבות מסויימות תורמת נוכחותם של פרטים הומוסקסואלים, כעוזרים ומסייעים בעבודות משק, לכשירותה של אוכלוסיית ציידים, יש לחפש אחר גנים הומוסקסואלים במאגר הגנטי של האוכלוסיה. שכן הבירור הטבעי רואה בהם גנים מועילים המגדילים את הכשירות הדארוויניסטית הכוללת של הפרטים הפוריים.
הבעייה בעקרון יסוד זה, שהוא מאפשר להתאים אינטרפרטציה הסתגלותית לכל התנהגות שמתחת לשמש. שירת הצפורים היא דוגמא מובהקת לכך.
כידוע, דעות החוקרים חלוקות באשר לתפקיד המדוייק של שירת הצפורים: האם תכליתה לציין בעלות על טריטוריה? האם מטרתה למשוך בנות זוג? האם היא כלי נשק במאבק הבין-מיני? או אולי שלושתם גם יחד? בשנים האחרונות צפה שאלה חדשה: מדוע יש צפורים ששירתן מורכבת וממושכת?
חוקרי הטבע נוהגים לחלק את קולות הצפורים לשני סוגים: קריאות קצרות ושירים מורכבים. תפקיד הקריאות הקצרות מוסכם על הכל: למסור על סוג הקורא, מינו, זהותו, מיקומו וכוי. אבל, אם את כל המידע הזה אפשר למסור ע״י קריאות קצרות ופשוטות – מה צורך יש בשירים ארוכים ומורכבים? יתירה מזאת, מאחר שהמנגנון השירי צורר הרבה אנרגיה וזמן, מה ראה הבירור הטבעי לשמרו ואף לשכללו? מחקר שנערך אך לאחרונה בקרב זמירי הסוף, ע״י קליב קצ׳פול, הצביע בוודאות על כך שהשיר הוא מכשיר של משיכה מינית. לראייה, משמוצא לו הזכר בת זוג הוא מזמר הרבה פחות (אצל זמיר הגומא נפסקת השירה כליל). אבל לא רק זאת, לעומת שיריהם הקצרים והפשוטים של צפורי שיר אחרות, שירת הזמיר עשוייה להמשך יותר מדקה תמימה ובאמתחתה רפרטואר של 50 שירים שונים המתבסס על מאגר של מאות הברות! מורכבות זאת מקנה לשירת הזמיר אותו מעמד אקסטראווגנטי שיש לזנב המפואר של הפאסיון. דהיינו – תוצאה מופלגת של מאבק בין-מיני עז. כדי לאמת תיזה זאת העדיף קצ׳פול שהשירה תהיה מורכבת וססגונית יותר בקרב מיני זמירים שהם פוליגיניים (מזדווגים עם מספר נקבות) מאשר בקרב המינים המונוגמיים. שכן נדרש מאמץ רב יותר למשוך מספר נקבות מאשר להקסים נקבה אחת. אלא שהממצאים, למרבה ההפתעה, הראו מגמה הפוכה: שיריהם של המינים הפוליגיניים היו הרבה יותר קצרים ופשוטים מאשר של עמיתיהם המונוגמיים. אך לא חלף זמן רב וקצ׳פול נחלץ מהמבוכה בהסבר נאות: מאחר שהזכר הפוליגיני ממהר לנטוש את זוגתו לאחר ההפרייה, ממילא אין חשיבות מרובה מבחינתה לאיכויותיו הזכריות. מה שחשוב בעיניה הוא גודל הטריטוריה שלו וכמות המזון שבה. במילים אחרות, מאחר שהיא מכירה את טיבו של ה״תכשיט״ שלה, אין לה עניין בסלסוליו אלא בתכל’ס.
לעומת זאת, אצל המינים המונוגמיים הטריטוריות קטנות יחסית והזכר נאלץ להשיג את מרבית המזון מחוץ לגבולותיה, אגב פשיטות ארוכות ונועזות בארץ לא נודעת. מכאן שנקבה נבונה תקפיד לבחור בזכר בעל איכויות משובחות של חוסן, אומץ והתמדה. הואיל וכך סלסוליו הארוכים והמסובכים של הזמיר באים לגולל באזני זמירתו את עלילות גבורתו, למען תעתר לו.6
הנה כי-כן, התאוריה הדארוויניסטית יכולה לספק הסבר מניח את הדעת לתופעה ספציפית ולהיפוכה גם יחד. במילים אחרות, במקרים רבים התיזות השונות לא ניתנות להפרכה חד-משמעית ע״י ניסוי ואימות תצפיתי. מטעם זה הגדיר פופר את הדארוויניזם בתאוריה פסוידו-מדעית, כלומר – תאוריה מטאפיסית, אף כי בעלת חשיבות ממדרגה ראשונה למדע. יתירה מזאת, לא זו בלבד שניתן להתאים אינטרפרטציה לכל התנהגות הסתגלותית, קשה גם להבחין בין התנהגות הסתגלותית שנרכשה באמצעות למידה ותרבות לבין התנהגות תורשתית. דוגמא לנושא זה אפשר למצוא במנהגם של קשישים בקרב קבוצות מסויימות של אסקימוסים.
כאשר מקורות המזון המתדלדלים מאלצים את השבט לתור אחר מושב חדש, נשארים הקשישים מרצונם במקום וממתינים למותם, כדי לא לעמוד בדרכו של השבט במסע הארוך והמפרך הצפוי לו. התאוריה של בירור שאירים מייחסת התנהגות זאת לקיומם של גנים אלטרואיסטים, המביאים פרטים קשישים ולא פוריים להקריב חייהם כדי לשפר את סיכויי ההשרדות של שאיריהם, נושאי הגנים שלהם. מעשה זה משפר את הכשירות הדארוויניסטית הכוללת של הקשישים ולפיכך זוכה לעידודו של הבירור הטבעי. אפשר, כמובן, להקיש ששבטי אסקימוסים אשר לא פיתחו גנים אלטרואיסטים כאלה ונטלו את זקניהם עימם, הצליחו פחות במאבק ההשרדות. אבל, ניתן גם לתת להתנהגות זאת הסבר תרבותי, בלי להזדקק כלל לגנים. אפשר לראות במעשה ההקרבה שבא לסייע לבני השבט, מנהג תרבותי ששכרו בצידו: מעשי ההקרבה של הקשישים זוכים להערכה ולתהילה, ושמותיהם תופשים מקום חשוב במסורת התרבותית של השבט.
הנה כי-כן, התודעה החדירה גורם לאבולוציה הביולוגית: אדם יכול לדבוק בשאיריו – לא בכוח הצווים הגנטיים, אלא מטעמי כבוד, תהילה, גאווה, הגשמה עצמית וכיו״ב ערכים מנטליים.
אי-יכולת זאת להבחין בין ביולוגיה לתרבות עומד ביסוד הפולמוס הנטוש בין חסידי הסביבה לבין הסוציוביולוגים, פולמוס שלא ניתן, ככל הנראה, להכרעה. תאודוזיוס דובז׳נסקי, מהדוברים הראשיים של זרם הסביבה, טוען כי הגנים האנושיים ויתרו על בכורתם כנושאי האבולוציה האנושית, לטובת סוכן חדש, לא ביולוגי: התרבות.
גם ז׳וליאן הקסלי שם את האצבע על ההבדל המהותי בין הפרוגרס הביולוגי הטהור, המניע את התפתחות בעלי החיים, לבין הפרוגרס האנושי. לטענתו, אנו נוטים לטשטש את ההבדל העקרוני בינינו לבין החיות. אנו נוטים לחשוב, אומר הקסלי, כי אם יש המשכיות בזמן בין עולם החיות לעולמנו, יש גם המשכיות של איכות. אבל, כשם שהאדים מהווים המשך חומרי של המים, אך שונים מהם בתכונותיהם, כך גם היחס בין החיות לבינינו. ההתפתחות אצל החיות היא לעולם סגורה ובסופו של דבר מוגבלת. לעומת זאת, ההתפתחות האינדיבידואלית של האדם אינה מוגבלת. היא נמשכת כל חייו ובכוחה לבור לה אין סוף כיוונים. האדם פיתח דרך חדשה של אבולוציה: העברה מאורגנת של הנסיון באמצעות מסורת. דרך זאת אינה כפופה לתהליך המכאניסטי של הבירור הטבעי, אשר חדל להיות נושאו הבלבדי של השינוי באבולוציה של האדם.
חידת יבלות הרגלים וביצי הזוחלים
אבל, בעיותיה של האבולוציה לא מתחילות באדם אלא מוקדם הרבה יותר, והן נוגעות במכאניזמים הבסיסיים והמרכזיים שלה.
כזכור נדחתה התיזה הלמארקיסטית, לפיה הלחץ הסביבתי גורם לאורגניזם לרכוש תכונות הנחוצות לקיומו ולהנחילן בתורשה. את מקומה תפשה התיזה הדארוויניסטית, לפיה מתחוללים השינויים באיברים ובתכונות באורח אקראי ועיוור, ורק אח״כ בא הבירור הטבעי ומשמר אותם שינויים המשפרים את כושר הסתגלותו של האורגניזם לסביבה. אלא שתיזה זאת אינה מספקת תשובה להופעתן של שכבות העור הקשות (יבלות) בכפות הרגלים של האדם, באחוריים של היען ובברכיו של הגמל. יבלות אלה נוצרות, כמובן, מתגובת תאי העור לשפשוף ולשימוש האינטנסיבי שהם נתונים בו. אבל כיצד להסביר את העובדה שהעור מתחיל להתקשות באיזורים אלה כבר אצל העובר, עוד טרם נתנסו בשפשוף כלשהו? התשובה הלמארקיסטית היא שיש כאן תכונה חיונית נרכשת של תאי העור המונחלת בתורשה, אך ״מהדוגמה המרכזית״ אנו למדים כי שום מידע אינו יכול לעבור מתאי העור לתאי הרבייה. מצד שני, ההסבר הדארוויניסטי, לפיו מוטציה מקרית יצרה יבלות דווקא באיזורים אלה, הוא כה בלתי מסתבר מבחינה סטטיסטית עד כדי לעורר אי-נוחות. אמנם נעשו נסיונות להציע הסברים בעלי אופי למארקיסטי ובסיס דארוויניסטי, כמו ״ההתפתחות המתועלת״ של וואדינגטון (מגדולי חוקרי התורשה בימינו), אך אי-הנוחות לגבי היבלות בעינה עתרה. אלא שאי-הנחת מהיבלות היא כאין וכאפס לעומת האצבע של דב הפאנדה וביצת הזוחלים, המדירות שינה מעיניהם של אבולוציוניסטים רבים.
לפאנדה הענקית יש אצבע שישית בגפיה הקדמיות, המאפשרת לה ללקוט ביעילות חוטרים של במבוק שהם מזונה העיקרי. האצבע השישית היא מקרה קלאסי של התמחות, היכול, כמובן, להיות הרה גורל לקיומה של הפאנדה אם יעלם הבמבוק מסביבתה. כך או כך הופעתה של אצבע נוספת כרוכה בפיתוחו של מערך חדש ומורכב של שרירים, עצבים, גידים ועצמות. על פי התורה הדארוויניסטית יכול מבנה כזה להיווצר בעקבות תהליך ממושך ומצטבר של מוטציות זעירות ואקראיות. אבל, וכאן הקושי30 לכאורה, אין בכל אחת מעשרות מוטציות הביניים הזעירות שום תועלת כלשעצמה, אלא בתוצאה הקולקטיבית הסופית. כיצד, אם-כן, התרחשו באופן אקראי, עצמאי ונפרד, ולא נכחדו ע״י הבירור הטבעי, כל אחת בתורה, במוטציות שליליות ומזיקות? הוא הדין בביצי הזוחלים.
כבוש היבשה ע״י החולייתנים החלה עם התפתחות הזוחלים מדו-חיים פרימיטיביים. אלא שהמעבר לחיי יבשה חייב שינוי קיצוני ודרמאטי בתכונות ביציהם. בהיותן חשופות לאויר הן היו צריכות לפתח קליפה שתמנע את איוד הנוזלים מתוכה. הן היו צריכות להיות מצויידות בחלמון ובאלבומין לאספקת מזון ונוזלים לעובר, בכיס אשפה פנימי לאגירת הפסולת של חילוף החומרים (בביצי היצורים הימיים הפסולת מופרשת הישר למים), בקרן עוברית לפיצוח הקליפה (העוברים הימיים מפרישים לצורר זה חומר כימי) וכד’. עכשיו, שינויים אלה, אם אינם מתואמים ואם אינם מתרחשים בעת ובעונה אחת, אין בהם תועלת. יתירה מזאת, כל אחד מהם בנפרד עלול להיות קטלני לעובר (כמו קליפה קשה ללא קרן לפיצוחה). אך להיות מתואמים מראש משמעו לייחס לאבולוציה מגמה ותכנית קבועות מראש, הפסולות מכל וכל ע״י האבולוציה הדארוויניסטית. מצד שני, להניח אפשרות שכל המוטציות הללו התרחשו בו-זמנית ובמקרה, היא בלתי אפשרית מבחינת ההסתברות הסטטיסטית. אם-כן, כלום נושאת האבולוציה בתוכה, ככלות הכל, מגמה (אורתוגנזיה) ותכלית (טלאולוגיה)? או, אולי, עלינו להשלים עם אמירתו של פישר כי ״הבירור הטבעי הוא מכאניזם ליצירת חוסר הסתברות ברמה גבוהה?״ אפשר, אמנם, הסבר שלישי והוא, שהמעבר מהחיים הימיים ליבשתיים התרחש באיטיות ובהדרגתיות בביצות שבשולי האגמים והימים, שם נוצרו במעורב תנאים יבשתיים וימיים גם יחד. כך או כך אין בהסבר כזה או אחר כדי לסלק כליל את סימני השאלה הרודפים אחרינו. אומר על כך ואדינגטון: להניח כי האבולוציה של מכאניזמים ביולוגיים מותאמים להפליא התבססה אך ורק על פעולת בירור מתור מאגר גדול של מוטציות, שכל אחת מהן נוצרה ע״י מקרה עיוור, משול למימוש ארכיטקטוני של בית ע״י זריקה אקראית של לבנים בערימה.
במילים אחרות, האם האבולוציה היא סדרה של תאונות מקריות, או שהיא פועלת במסגרת חוקיות בעלת מגמה פנימית?
האבולוציה – סדרת תאונות מקריות?
כידוע, זה אלפי שנים שהאדם למד כיצד להתערב באבולוציה ולהאיץ את מהלכה האיטי באמצעות הכלאות מלאכותיות של זני צמחים ומיני בעלי חיים. הצלחנו לפתח גזעי בקר שופעי חלב ומשופעי בשר ולהניב זני פירות בשריים וגדולים. אעפי״כ לא הצלחנו, וככל הנראה לא נצליח, לפתח שזיף בגודל של אשכולית, או להפוך חזיר לסוס. איננו יודעים מדוע, אלא זאת שתכונות אלה חורגות אל-נכון מהפוטנציאל האבולוציוני של השזיף ושל החזיר.
יהיה משמעותו של מונח זה – הפוטנציאל האבולוציוני – אשר יהיה, עובדה היא שלא ניתן להגביר ולחזק תכונותיו של מין כלשהו מעבר לגבול מסויים.
בניסוי נודע שנערך בזבובי התסיסה המפורסמים (Drosophila), הצליחו ע״י הכלאות חוזרות ונשנות, במהלך 20 דורות בלבד, להגריל את מספר זיפיהם מ-36 ל-56, אך לא מעבר לזה. כאשר הגיעו הזבובים ל-56 זיפים הם הפכו עקרים ותמותתם גדלה בשעור ניכר.
אם בזבובי תסיסה עסקינן, הבה נזכיר תופעה מענינת אולי אף יותר, אשר השלכותיה לא ברורות ואפשר אף מביכות למדי.
כאשר שני גנים רצסיביים מסויימים של הדרוסופילה נפגשים ויוצרים באחד הצאצאים גן רצסיבי הומוזיגוטי, נולד צאצא זה ללא עיניים. עכשיו, אם מכליאים את הצאצאים נטולי העיניים אלה עם אלה, הרי כעבור מספר דורות שבים להיוולד זבובים נורמליים בעלי עיניים תקינות.
כיצד אפשרי הדבר? אין, כמובן, להעלות על הדעת שבמרוצת דורות ספורים התפתחה מחדש עין באותו תהליך מקרי ואקראי שארך בטבע מאות מיליוני שנים. כיצד, אם-כן? האם המידע ליצירת העין היה אצור בגנים כפולים המצויים כרגיל במצב רדום? אבל אם כך מדוע לא שבים האפונים הקמוטים של מנדל לחלקות הפנים של הוריהם ההטרוזיגוטים? או שמא הותוותה יצירת העין ע״י תכנית-אב לתיקון פגמים, הפועלת ברמה גבוהה יותר מאשר הגנים? זאת ועוד, העיניים המחודשות של צאצאי הזבובים העוורים והעיניים של הזבובים הנורמליים, הן איברים הומולוגיים (איברים של אורגניזמים שונים, אך בעלי דמיון מבני רב), למרות שנוצרו מקומבינציה שונה של גנים.
מה משמעות הדבר? העובדה שהתכנית המבנית של ידי האדם (העצמות, השרירים, העצבים וכלי הדם) דומה דמיון רב לגפיים הקדמיות של כלב, לכנף הצפור ולסנפיר הלווייתן, למרות שתיפקוד איבריהם שונה, נובעת – עפ״י הדעה הרווחת – מכך שהמערך הגנטי הנושא את תכנית הבנייה של איברים אלה הונחל ממין למין, ממשפחה למשפחה, מסדרה לסדרה, ממחלקה למחלקה, וכיו״ב, ללא שינוי, מאחר שהשיג מבניות מיטבית המתאימה לצורות תפקוד שונות. אבל, אם העיניים של זבוב התסיסה אכן נוצרו ממערך שונה של גנים, אזי את התשובה להומולוגיה יש לחפש במקום אחר. כלומר, אפשר וביסוד האבולוציה קיימת תכנית-אב המאפשרת לפתח אין-סוף ואריאציות של תכונות וצורות, אך במסגרת מספר מוגבל של מוטיבים בסיסיים. ואריאציות אלה, כפי שהראה ד’ארסי תומפסון ב-1917, פועלות במסגרת חוקיות גאומטרית-מתימטית מובהקת. תומפסון צייר בעל חיים על סריג של קואורדינטות וע״י עיוות גאומטרי מסויים של הסריג, הניתן לביטוי מתימטי, הצליח לגזור דג קיפוד מהתכנית המבנית של דג שמש, וגולגולת אדם מגולגולת של באבון ושימפנזה. האם יש בכך כדי לחזק את ההנחה בדבר תכנית-אב פנימית לאבולוציה?
למותר לציין כי מסקנות אלה אינן מקובלות על מרבית הביולוגיים, ולו מהסיבה שמונחים כמו תכנית-אב, מגמה, תכלית וכיו״ב מחדירים למטריאליזם הדארוויניסטי יסודות מטאפיסיים חשודים. כך אירע למשל לחוק ה״חזרה״ של ארנסט האקל (1919-1834), ביולוג ופילוסוף גרמני, לפיו עובר הפרט במהלך האונטוגנזה שלו (התפתחות הפרט מהביצית המופרית לבגרות) דרך שלבי האבולוציה שעברו המינים שקדמו לו. ובניסוחו של האקל: האונטוגנזה היא חזרה על הפילוגנזה7 .
כבר קודמיו ציינו את העובדה המעניינת, כי שלבי ההתפתחות הראשונים של עוברי יונקים דומים31 לצורות ההתחליות של צפורים, זוחלים ודגים. דוגמא מובהקת לכך הם נוכחותם של כיסי זימים בעוברים של צפורים ויונקים. אך בניגוד להאקל הם הדגישו את העובדה שהדמיון בין הצורות מתקיים בין השלבים העובריים ההתחלתיים של המינים השונים ולא בינם לבין הצורות הבוגרות שלהם. למשל, העוברים והוולדות של הקופים דומים הרבה יותר לתינוק האנושי מאשר לקוף המבוגר. במילים אחרות, בתהליך האונטוגנזה מופיעים תחילה קוי המבנה המשותפים למערכת, אח״כ למחלקה, לסדרה, למשפחה ולסוג, ורק בשלב מאוחר יותר עוברים העוברים התמיינות לפי המאפיינים הספציפיים של כל מין. אף שחוק החזרה של האקל נדחה ע״י הביולוגים, ואף כי אין בדמיון הצורני בין מיני העוברים השונים כדי ראיה לקיומה של תכנית-אב כלשהי, אין הסבר מניח את הדעת להופעתם של כיסי זימים בעוברים אנושיים. מדוע שמרה האבולוציה על צורות קדומות אלה ומה חשיבותן להתפתחות היונק? האם זו נטייתה השמרנית לשמר צורות ומבנים מוצלחים, כל עוד אינם מפריעים, או אולי יש כאן ביטוי נוסף של עקרון ההומולוגיה? הבה נסיים הרהורים אלה בדבר קיומה האפשרי של תכנית-אב לאבולוציה, בפרשה המרתקת והתמוהה של הכיסנאים (מרסופיאלים) החיים באוסטרליה ובדרום אמריקה.
חידת המרסופיאלים
לפני 65 מיליון שנה לערך, זמן קצר לפני ההסתעפות הגדולה של היונקים, ניתקו אוסטרליה ודרום אמריקה מיבשת אפריקה ונותרו בבידוד מוחלט למשך עשרות מיליוני שנים. בפרק זמן זה התרבו היונקים בהצלחה רבה ממין חדף קדמון והסתעפו לסדרות, למשפחות ולמינים מגוונים. והנה, הפלא ופלא, למרות שלא היה שום מגע בין אפריקה והגוש האירו-אסיאתי מזה, לבין אוסטרליה, ניו-זילנד ודרום אמריקה מזה, התפתחו בשני גושי היבשות אותם מינים כמעט, בהבדל אחד: בגוש הראשון התפתחו יונקים שלייתיים, ואילו בגוש השני התפתחו יונקים כיסנאים. בצפון אמריקה ובדרומה, שהיו מנותקות זו מזו עד להיווצרות הגשר היבשתי ביניהם לפני 3 מיליון שנה לערך8 , התפתחו באורח עצמאי ובנפרד, מינים מקבילים. כנגד הטורפים השלייתיים של הצפון התפתחו טורפים כיסנאים בדרום: נמרים, כלבים, חתולים, צבועים, זאבים ועוד. בין אוכלי העשב התפתחו בדרום צורות מקבילות (שלייתיות) של גמלים, סוסים וקרנפים.
השאלה הנשאלת היא, כמובן, הכיצד? אם האבולוציה היא תהליך עיוור, המתקדם באמצעות מוטציות אקראיות, ואם לא התקיים כל מגע בין היבשות, מדוע לא התפתחו בדרום אמריקה ובאוסטרליה מפלצות בעלות שלוש עיניים, ארבעה זנבות ושש רגליים? כיצד אפשר שמתוך מגוון אין-סופי של אפשרויות התפתחו דווקא צורות חיים מקבילות (להוציא הקנגורו, דובי קואלה ומינים ספורים אחרים) לאלו שנוצרו בנפרד ובאורח עצמאי באפריקה, באסיה ובאירופה? ההסתברות הסטטיסטית להיווצרות מיקרית של דמיון כזה היא אפסית. מהי, אם-כן, התשובה? האם האורגניזמים שנוצרו במהלך האבולוציה מהווים את צורות החיים המיטביות ובעלות כושר ההסתגלות הגבוה ביותר לתנאי הסביבה השוררים על-פני כדור הארץ, ולפיכך רק צורות אלה נשרדות? או שמא קיימת תכנית-אב פילוגנטית – הקובעת מראש אלו ואריאציות יכולות צורות החיים השונות לפתח?
היווצרות המינים: מפלצות בנות מזל?
מצדדי ״הסינתזה המודרנית״ של ז׳וליאן הקסלי, המייצגים את הזרם המרכזי של קהיליית המדענים, דוחים על הסף קיומה של תכנית-אב. לטענתם מסוגלת האבולוציה לפתח אורגניזמים בכל הצורות, הגדלים והמבנים. הסיבה שלא פיתחה פרה בעלת שני ראשים, קדמי ואחורי, היא שאין גומחה מתאימה לצורה זאת של חיים. אחרים מכירים במגבלות לאפשרויות המורפולוגיות, הנובעות מהתכונות המכאניות של אבני הבניין של החיים. לטענתם, הסיבה שלחיות היבשה יש ארבע רגליים היא שזוהי צורת ההליכה המיטבית ביבשה. עניין זה מביא אותנו לסוגיה אחרת, שנוייה אף היא במחלוקת, בדבר היווצרות מינים חדשים מתוך הקיימים.
על-פי ״הסינתזה המודרנית״, השלטת במחקר האבולוציה זה 40 שנה, מתנהלת האבולוציה באיטיות רבה (בתנאי סביבה יציבים), באמצעות שינויים זעירים המצטברים והולכים במשך מיליוני שנים. מינים חדשים נוצרים בהשפעת בידוד גאוגרפי והיווצרות גומחות סביבתיות חדשות, היוצרות לחץ על הבירור הטבעי להעדיף מוטציות מסויימות בעלות התאמה גבוהה יותר לסביבה מאשר המערך הגנטי של המין המקורי. הקושי בהסבר זה נעוץ במנגנון האיטי שלו, אפילו במושגי הזמן האדירים של האבולוציה. עפ״י חישוביו של הולדין נדרשים 300 דורות כדי להחליף אלל אחד במשנהו באתר כלשהו ע״פ הכרומוסומים. ומאחר שמינים מסויימים נבדלים באללים שלהם ב-1000 אתרים שונים, נדרשים 300,000 דורות כדי לפתח מין חדש. זאת ועוד, התיזה של השתלשלות המינים באמצעות צעדים זעירים אינה נתמכת לכל אורך הדרך ע״י הממצאים הפלאונטולוגים.
שרידי העצמות והמאובנים אינם מצביעים על מעבר הדרגתי וחלק מצורות מורפולוגיות ישנות לחדשות, אלא על התמדה ממושכת של המין בצורתו הקיימת, עד לשינוי חד ופתאומי המיוצג ע״י המין החדש. הנסיון המסורתי לתלות פערים פלאונטולוגים אלה באופיו המתכלה של החומר האורגני ובנסיבות אקלימיות ואחרות, המונעות מחוליות ביניים רבות להותיר מאובנים בעקבותיהם – אינו מספק רבים וטובים.
העדרן של חוליות ביניים רבות מעלות אצל חוקרים לא מעטים את המחשבה שמא לא היו ולא נבראו. אפשר, גורסים אלה, שהאבולוציה אינה מתקדמת עקב בצד אגודל, אלא בצעדים גדולים ונחשונים. היה זה ריצ׳רד גולדשמידט, אשר הציע ב-1930 לראות במאקרו-מוטציות את המנגנון המרכזי של התפתחות המינים. דהיינו, האבולוציה מתקדמת בדילוגים, באמצעות ״מוטציות של המערכת״ (Systemic Mutation) ולא באמצעות חוליות עוקבות, הנבדלות זו מזו בשוני זעיר.
הקושי שניצב לפני גולדשמידט היה להסביר, בלי להזדקק למעשי ניסים, כיצר מתרחש דילוג כזה וכיצד נמצא תואם לתנאים הסביבתיים. שכן, כל עוד מדובר בשרשרת ארוכה של מיקרו-מוטציות, אזי כל שינוי קטן נבחן עפ״י מידת התאמתו לסביבה: אם הוא משפר את כשירותו הדארוויניסטית של האורגניזם – יישרד, אם גורע – ייכחד. אבל אם מדובר במאקרו-מוטציה, הרי על-מנת שכל השינויים הזעירים הכלולים בה יפעלו בהארמוניה אלה עם אלה וגם יתאימו לסביבה, דרוש לא פחות מנס. תשובתו של גולדשמידט: ״מפלצות בנות המזל״.
סוד גלוי הוא, טען גולדשמידט, שאחת לכמה זמן נולדים להורים נורמליים יצורים מפלצתיים. רובן המכריע של המפלצות אלה אינן מאריכות ימים, אך אפשר שאחת לתקופה ארוכה נולדת מפלצת, השונה באורח משמעותי מהוריה; ועשוי לקרות ששוני זה יתאם ביעילות יתירה את הסביבה הקיימת או את הסביבה העומדת בשלב השתנות קיצונית. מה שברור הוא, שאם הצליחה מפלצת כזאת להשרד ולעמוד במאבק הקיום, היא עשוייה להוות מקור ומוצא למין חדש.
במרוצת הזמן ידאג הבירור הטבעי לשפץ את איבריהם ואת תכונותיהם של צאצאיה, עד שיביאם להתאמה מלאה עם הסביבה. לדוגמא, אחת השאלות הקשות הניצבות לפני האבולוציה היא איר נוצרו הנוצות בקרב הצפורים. ברור לכל שזהו איבר מורכב מכדי שיתהווה ע״י מוטציה אחת. מצד שני, איזו תועלת יכלה להיות לכל שלב משלבי הביניים הדרושים למעבר מקשקשים של זוחל לנוצות של העוף, עבור דורי-דורות של אבות הצפורים? לדעת פרופ’ דן כהן מהאוניברסיטה העברית בירושלים, אין קושי להניח היווצרות עצות מהתארכות והתפצלות הדרגתית של קשקשים. מיינרד סמית ואחרים גורסים ששלבי הביניים סייעו לדו-חיים בביצה במלאכת הנשימה – הרבה לפני שהפכו יצורי יבשה. הגם שהסברים אלה אינם מפריכים את אפשרותן של מאקרו-מוטציות, דחתה הביולוגיה המודרנית את תורתו של גולדשמידט, אך יצרה במקומה גרסה משופרת, הקרויה בפי הוגיה – גולד ואלדרירז׳ – ״שיווי משקל מפוסק״. האבולוציה, מסכימים השניים, אינה מתנהלת בצורה קווית והדרגתית. למשך זמן מה מתמיד האורגניזם בצורתו, ואז מתרחשת צמיחה מואצת ומסועפת, בדומה להנצת עץ בעקבות גיזום. כל אותם ניצנים הנושאים יתרונות מפורשים (כמו הקטנת מספר האצבעות והגדלת מימדי הגוף אצל הסוס), זוכים לעידודו של הבירור הטבעי ומביאים בסופו של דבר להתבססות מהירה של מינים חדשים.
תיזה אחרת מייחסת את הכח המניע של השינויים לא לגורמים סביבתיים-חיצוניים, אלא לפנימיים. ניקח, לדוגמא, שני טיפוסי חיות. האחד ספציאליסט, המבסס קיומו על מזון מאוד מסויים שגדל בסביבה ספציפית (כמו דב הקואלה האוסטרלי, הניזון מסוג מסויים של עלי אקליפטוס. הוא מצוייד במערכת עיכול המנטרלת את הרעל של שמן האקליפטוס וכן טפרים חדים המאפשרים לו לטפס ביעילות על גזעי העץ החלקים), ואילו השני ג’נרליסט, שתפריטו32 מורכב מסל עשיר של עלים ועשבים והיכול להתאים עצמו לסביבות שונות. התוצאה: מהספציאליסס יתפתחו הרבה יותר מינים מאשר מהג’נרליסט. הסיבה פשוטה. הספציאליסטים תופשים ערוגה ביולוגית צרה ולפיכך אינם מופרעים ואינם מוטרדים מהתהוותם של מינים קרובים, כאשר כל אחד מהם תופש ערוגה נפרדת. הג’נרליסט, כנגד זאת, מזונו מגוון ולפיכך מועד לתחרות ולהתנגשויות אין-ספור מצד מינים ג’נרליסטים כמוהו. לפיכך אינו סובל מינים קרובים לו בשכנותו. הג’נרליסט, איפוא, הוא בעל כושר הסתגלות לשינויים ולתנאי סביבה מגוונים, לכן גם בעל כושר השרדות גבוה. הספציאליסט, לעומתו, סובלן מאין כמוהו, אך משום התמחותו הצרה הוא צפוי להכחדה עם כל שינוי משמעותי המתרחש בסביבתו.
במילים אחרות, המרכיב העיקרי של השינוי, לפי תיזה זאת, הוא גורם פנימי: הספציפיות היתירה.
שאלות פתוחות ותמיהות אחדות
הינה כי-כן, האבולוציה הדארוויניסטית היא המנגנון הבלבדי והחשוב ביותר שהוצע מעולם למתן הסבר רציונלי להשתלשלותם ולהתפתחותם של בעלי חיים – על איבריהם ותכונותיהם. אך עובדת היותה דיסציפלינה הסטורית באופיה, המפרשת ממצאים וארועים לאחר התרחשותם, אינה מאפשרת לה, ככלל, לאמת את הנחותיה באופן אמפירי ובכך נשלל ממנה המעמד המדעי המובהק שכה נחוץ לה.
נקודת התורפה העיקרית של ההסבר האבולוציוני בכך, שאנו יודעים מראש כיצד נראות, למשל, פרסות הסוסים, לפני שאנו שואלים מדוע התפתחו כפי שהתפתחו ומה תועלתן. לשון אחר, במקום לטעון טענות א-פריורי, כנהוג במדע, עושה ההסבר האבולוציוני שימוש בטענות פוסטריורי. ומאחר שהאבולוציה עוסקת בדיעבד, כמעט ואי-אפשר להכריע בין תאוריות חליפיות בשיטת ההפרכה.
דוגמא נאה לכך היא האלטרואיזם של הנמייה הננסית. לנקבות בקרב יצורים אלה מעמד עדיף על הזכרים. כאשר אחד הזכרים חולה, הוא זוכה לטיפול מסור של הנקבות ומקבל גישה מועדפת למזון. ההסבר להתנהגות זאת לא נעוץ בעקרון של בירור שאירים, אלא באופי המזון של הנמיות הננסיות. מאחר שהן ניזונות מחרקים המצויים בשפע והניצודים באורח אינדיבידואלי, הפרטים החולים אינם מכבידים על אוכלוסיית הנמיות באותה מידה שהם מכבידים על אוכלוסיה של חיות ציד. להיפך, ככל שאוכלוסיית הנמיות תגדל כן יגדל בטחונו של כל פרט בקרבה מפני טורפים בכח.לכן, יש אינטרס ברור לקבוצה להבריא את פרטיה. הבעיה עם הסבר זה שקשה לאמתו או להפריכו. יתירה מזאת, על פי תיזה זאת האינטרס של חיות הציד הוא להניח לחולים למות. אולם, מסקנה זאת אינה עולה בקנה אחד עם דאגתם ומסירותם של כלבי הפרא האפריקנים לעמיתיהם החולים והקשישים.
תמיהות רבות מעין אלה עולות ומבצבצות בעקבות ההסברים הסוציוביולוגיים. אם ההתנהגות האלטרואיסטית של כלבי הפרא האפריקנים מוסברת ע״י העקרון של בירור שאירים, מדוע אינו חל על יחס האריות לגוריהם? מדוע מגרש האריה את גוריו הבוגרים ומסכן בכך מנת גנים עצמיים השווה ואף עולה על מה שהוא נושא בגופו הוא? זאת ועוד, אריה המשתלט על עדה חדשה הורג את הגורים הזכרים וגורם ללביאות להפיל את עובריהן. מעשה זה משרת, ללא ספק, את האינטרס של האריה, אך מנקודת מבטה של הלביאה יש בכך אובדן מלא של השקעתה. ומאחר שאין להחשיד את הבירור הטבעי בשוביניזם זכרי, מדוע התעלם כליל מהאינטרס של הלביאות? מדוע אינן נלחמות עם האריה החדש ומגרשות אותו? מדוע הן לא אימצו את המבנה המטריארכלי של הפילות, המגביל את הזכרים לתפקיד ההרבעה בלבד? או לחילופין, מדוע לא אימצו אורח חיים סוליטרי כשל הנמרות, המגדלות בכוחות עצמן את גוריהן ומבטיחות בכך את השקעתן מירידה לטמיון?
פרופ’ דן כהן גורס כי במצב של ניגוד אינטרסים, צד אחד מנצח, ובמקרה הספציפי הזה ידם של האריות היתה על העליונה. ד״ר אבי שמידע, אף הוא מהאוניברסיטה העברית, מייחס זאת לגישה הפלורליסטית של הטבע, המכריע בין פתרונות שונים באמצעות ה-ESS.
לפרופ’ אמוץ זהבי גישה שונה לחלוטין. לדעתו, התנהגות האריה כדאית גם ללביאה, שכן אילו היה האריה מניח לה לסיים את הריונה ורק אז טורף את גוריה, היה הפסדה מלא. העובדה שהיא מפילה תיכף להשתלטות האריה על העדה, חוסכת לה חודשים של הריון סרק.
דומה כי תשובה זאת אינה נקייה מבעייתיות. שכן, יש להוכיח שהאריות היו אכן טורפים את גורי הלביאות גם לאחר שהיו שוהים במחיצתן בכל תקופת הריונן. זאת ועוד, אף שהן מרוויחות חודשים של הריון סרק, ההשקעה שכבר השקיעו בעובר – יורדת בכל זאת לטמיון. לפיכך גם עפ״י הסבר זה היו יוצאות נשכרות יותר אילו אימצו, למשל, את אורח חייה של הנמרה.
אך הבה נמשיך במסע תמיהותינו. אפשר להסביר את מנהגן של קופות לאמץ יתומים בדרגת הקירבה הגנטית הקיימת בין פרטי הקבוצה, אבל יש קופות הנוהגות גם לגנוב תינוקות מאימותיהם ולטפל בהם כאילו היו צאצאיהן. התנהגות זאת תמוהה, שכן אינה תורמת דבר לכשירות הדארוויניסטית הכוללת שלהן, יתירה מזאת, היא בזבוז זמן והשחתת אנרגיה על גנים לא להן.
ומהקופים להומינידים. המסתורין הגדול של הופעת האדם והגידול המדהים בנפח מוחו, כרוך בשתי שאלות: מהם הגורמים הסביבתיים שהביאו אותם לפתח הסתגלות שונה מאשר הקופים, ומה גרם להם להגיע כה רחוק?
כרגיל רואים בנטישת היער, במעבר לסוואנה הפתוחה ובשינוי התפריט התזונתי, את הגורמים העומדים מאחורי ההליכה על שתי רגלים, וכתוצאה מכך לשחרור הידיים ולהתפתחותו של המוח. אך מהלך דומה עשו גם הבאבונים, מדוע לא צעדו בעקבות האדם?
זאת ועוד, קופים רבים עושים שימוש מיומן ומתוחכם בידיהם. שימפנזות, גורילות ואורנגוטנגים מסוגלים להלך על גפיהם האחוריות. מדוע לא הוסיפו כישורים אלה להשתכלל? מדוע לא הביאו בהתאם לגידול של המוח? ועוד, יש המייחסים לקשר המונוגמי ולמבנה המשפחה של האדם תפקיד חשוב בפיתוח הכושרים האנושיים היחודיים. עכשיו, הן הקשר המונוגמי והן המבנה המשפחתי התאפשרו, לדעת חוקרים, בזכות אובדן הייחום העונתי (המאפיין את ממלכת החי), דבר המאפשר לנקבה האנושית להיות זמינה מבחינה מינית כל ימות השנה ולהדק בכך את הקשר המשפחתי. והנה, מסתבר שגם האורנגוטנגים מזדווגים כל ימות השנה, בלי להתחשב בייחומה של הנקבה, ובכל זאת נותר האורנגוטנג אחד היצורים היותר בודדים בטבע.
באשר לסוסים, טוענים כי עם פיתוח הפרסה והגידול בגוף הגיעו לנקודה מיטבית שאותה אין לשפר יותר ע״י מוטציות. אך כלום הזברה לא היתה יוצאת נשכרת אילו הצמיחה קרניים או ניבים חדים שהיו מאפשרים לה לעמוד ביתר הצלחה מול הטורפים? ומה באשר לתוספת מוח ותבונה, כלום היתה ניזוקה מכך? או אפילו פחות מזה. הסוס אינו מסוגל למזג את חוש המישוש והראייה באותו אופן שהקוף והאדם עושים זאת. לגבי דידו עצם לצידי הדרך הוא פשוט גוף לא מוכר.
מטעם זה הוא עשוי להבהל למראה אבן. האין במיזוג זה שיפור ויתרון לגביו? ומכאן לצפורים. הקוקיה מצאה דרך אידאלית להפצת הגנים שלה בלי33 להתאמץ. אך הצלחתה פירושה כשלון מוחלט של הפונדקאית. מסכנה זאת לא רק מאבדת את גוזליה, כי אם גם מכלה כוחותיה כדי לפרנס את גוזלי הקוקיה שרעבונם אינו יודע שובע. הכיצד לא דאג הבירור הטבעי לפתח בפונדקאיות מנגנוני זהוי והשמדה של הביצים הזרות?
איר זה שמנגנון כה משוכלל ליצירת מיטביות, כמו הבירור הטבעי, המשפר התנהגויות של בעלי חיים עד לדיוק מתימטי, לא הכחיד צפורים שאינן מסוגלות להבחין בין גוזליהן לגוזלים המפלצתיים של הקוקיה?
ומעניין לעניין באותו עניין. גמל-שלמה מסתכן במוות כל אימת שהוא בא לממש את מצוות פרו ורבו. עפ״י הסוציוביולוגיה, גורלו כפרט אינו חשוב – כל עוד הוא מצליח להפיץ את הגנים שלו. להיפר, בשמשו טרף לנקבה הוא מספק לה מנת חלבונים עשירה המשפרת את סכויי ההשרדות של צאצאיו. אך אם הגיון זה נכון לגבי גמלי-שלמה, עכבישים ויבחושים (מסויימים), מדוע אינו חל על כל הזכרים בטבע? ומצד שני, בשמשו טרף לנקבה המופרית מאבד הזכר סיכוי להפיץ את הגנים שלו בעשרות נקבות נוספות. מדוע, אם-כן, לא צייד אותו הבירור הטבעי במנגנוני הגנה והתחמקות מהנקבה הטורפנית? פרופ׳ זהבי טוען, כי אם סיכוייו להזדווג עם נקבות נוספות ממילא קלושים, מוטב לו להאכל. אך מדוע יהיו סיכוייו להזדווג בפעם שניה ושלישית קלושים יותר מאשר בפעם הראשונה? הוא הדין לגבי הקניבליזם הרווח בקרב מינים לא מעטים. אם תועלתו של הקניבליזם לכשירותם הדארוויניסטית של דגים, נמלים וחזירים היא כה מובהקת, מדוע אין הקניבליזם חזיון נפרץ בטבע? ולבסוף, אם בעלי החיים, על מיניהם השונים, הם רק כלי תחבורה של הגנים, ואם הקרבת החיים למען שלושה צאצאים הוא מעשה נכון ונבון מבחינת הגנים, מדוע לא כל בעלי החיים מגלים נכונות כזאת להקריב את עצמם?
שאלות אלה ואחרות זכו לתשובות רבות ושונות. אך כלום נאמרה, ולו בתחום אחד, המילה האחרונה? אל-נכון לא. אפשר שנמצא כי מכל ספרי הדעת הנכתבים והולכים בידי האדם, ספר הטבע הוא המופלא והמורכב שבהם, ולו מהטעם האחד שחידת ההוויה האנושית קשורה באלפי נימים גלויים ונסתרים בעקרונות המעצבים והמנחים את תכונותיהם והתנהגותם של יצורים חיים אחרים, לרבות הפחותים שבהם. ■

שעורי קירבה אלה מוצגים, כמובן, בצורה סכימטית. במקרים רבים מופרית המלכה ע״י מספר זכרים. הואיל וכך הסיכוי של אב משותף לשתי פועלות הוא 1 חלקי n, כאשר n הוא מספר הזכרים שהפרו את המלכה. ↩
תורת המשחקים נוסחה ב-1940 ע״י פון ניומן ואוסקר מורגנשטרן בספרם ״תורת המשחקים והתנהגות כלכלית״, במטרה לנתח קונפליקטים אנושיים בחיי יום-יום – כמו ניגודי האינטרסים הקיימים בין קונה למוכר בחנות – ובמצבי מלחמה. התורה מצביעה על האסטרטגיה האופטימלית שיש לאמץ במצבים של ניגוד אינטרסים. ↩
לפי וין-אדווארדס, העובדה שרק 4% מהזכרים בעדר מרביעים כ-80% מהנקבות, פועלת להגבלת הילודה של האוכלוסיה. קשה להבין את הגיונו של וין-אדווארדס, שהרי מה שקובע את שיעור הילודה הוא מספר הנקבות המעוברות ולא מספר הזכרים שהרביעו אותן. ↩
האנוכיות באה להוות משקל עד לאלטרואיזם, המתאר תופעות של עזרה לזולת בקרב בעלי חיים. שכן, מנקודת המבט של הגנים, ההורה המסכן חייו למען שאיריו אינו עושה מעשה אלטרואיסטי אלא אנוכי; הוא מקריב כמות קטנה של גנים שלו תמורת כמות גדולה יותר של העתקיהם השמורה אצל שאיריו. ↩
כל תאור התפתחותם של ההומינידים הוא בגדר השערה, המתחזקת או משתנה עם כל ממצא פלאונטולוגי חשוב חדש. ככל הנראה, לעולם לא נוכל לקבוע בוודאות אם שלד זה או אחר שנגלה מייצג את קוף האדם האחרון או את האדם-קוף הראשון, שכן אין ברשותנו קריטריון חד וברור להגדרת האדם. אם יצירת כלים, הליכה זקופה, מבנה שיניים ולסת, גודל מוח מעל נפח מסויים וכיו״ב, מהווים קריטריון ליצור האנושי – מסוגלת הפלאונטולוגיה לקבוע (ככל שתגלה ממצאים נוספים) מי היה האדם הראשון. אבל, אם הקריטריון לאנושיות הוא בכושרים פסיכיים, כמו שפה, תודעה וחשיבה, אשר אינם מותירים אחריהם מאובנים, לא נגיע לעולם להכרעה בסוגיה זאת. ↩
ממחקרו של ד״ר קן יאסוקאווה מאוניברסיטת רוקפלר, שהתפרסם לאחרונה, נמצא חיזוק לסברה שמגוון השירים ברפרטואר נועד ליצור רושם כאילו הטריטוריה מאוכלסת ומוגנת ע״י צפורים. דהיינו, כל אימת שהצפור משנה מקומה ועוברת לענף חדש היא מזמרת שיר שונה, כאילו היתה צפור אחרת. שיטה זאת מזכירה את טכניקת ההגנה של לוחם בודד המתרוצץ בין העמדות הריקות ויורה מכל אחת מהן כדי להטעות את התוקפים וליצור אצלם רושם שכל העמדות מאויישות ע״י לוחמים. ↩
ממלכת החי מתחלקת ל-26 פילות. היונקים, למשל, משתייכים לפילה של בעלי חוט שדרה, הכוללת את הדגים, הדו-חיים, הזוחלים והצפורים. ↩
היווצרות הגשר היבשתי חוללה את הטרגדיה הביולוגית החמורה ביותר של העת המודרנית. החיות השלייתיות של הצפון, אשר נדדו דרומה, דחקו את רגלי עמיתיהן הכיסנאיות והביאו להכחדתן. נראה כי ריבוי מיני היונקים בגוש האפריקני והאירו-אסיאתי, יצרו תנאי תחרות קשים יותר ולפיכך פיתחו בעזרת הבירור הטבעי מנגנונים יעילים יותר מאשר הכיסנאים. ↩
